

Oligoarginine peptides slow strand annealing and assist non-enzymatic RNA replication
- PMID: 27657866
- PMCID: PMC5061144
- DOI: 10.1038/nchem.2551
Oligoarginine peptides slow strand annealing and assist non-enzymatic RNA replication
Retraction in
-
Retraction: Oligoarginine peptides slow strand annealing and assist non-enzymatic RNA replication.Nat Chem. 2017 Nov 23;9(12):1286. doi: 10.1038/nchem.2885. Nat Chem. 2017. PMID: 29168488 Free PMC article.
Abstract
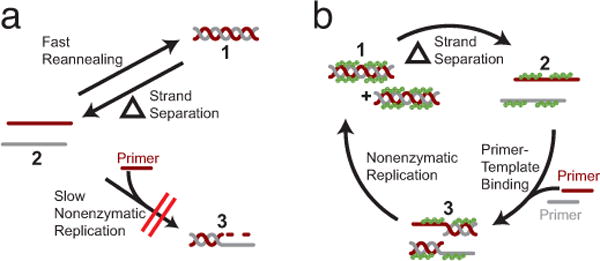
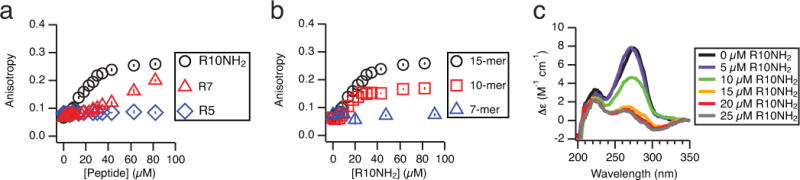
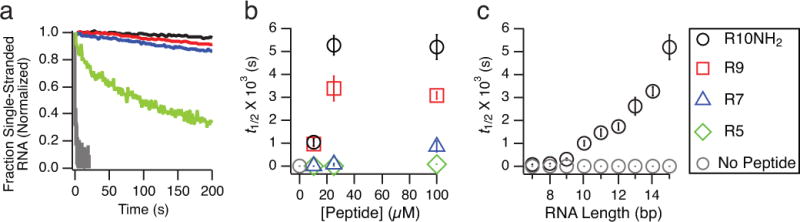
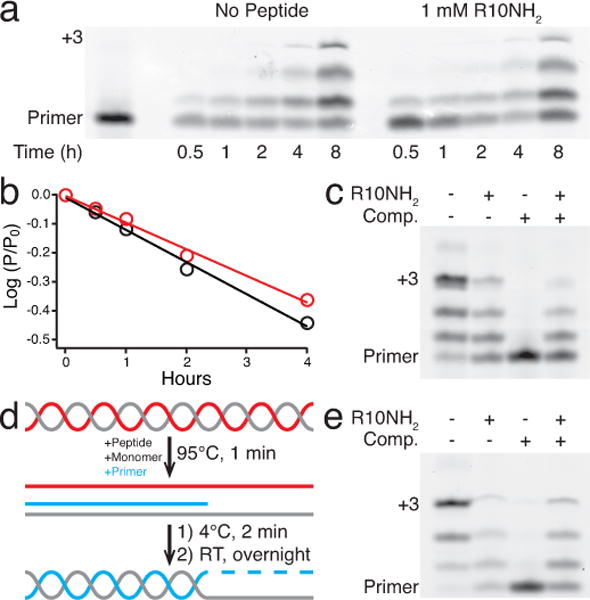
The non-enzymatic replication of RNA is thought to have been a critical process required for the origin of life. One unsolved difficulty with non-enzymatic RNA replication is that template-directed copying of RNA results in a double-stranded product. After strand separation, rapid strand reannealing outcompetes slow non-enzymatic template copying, which renders multiple rounds of RNA replication impossible. Here we show that oligoarginine peptides slow the annealing of complementary oligoribonucleotides by up to several thousand-fold; however, short primers and activated monomers can still bind to template strands, and template-directed primer extension can still occur, all within a phase-separated condensed state, or coacervate. Furthermore, we show that within this phase, partial template copying occurs even in the presence of full-length complementary strands. This method to enable further rounds of replication suggests one mechanism by which short non-coded peptides could have enhanced early cellular fitness, and potentially explains how longer coded peptides, that is, proteins, came to prominence in modern biology.
Conflict of interest statement
No competing financial interests are declared.
Figures
Similar articles
-
Rolling-circle and strand-displacement mechanisms for non-enzymatic RNA replication at the time of the origin of life.J Theor Biol. 2021 Oct 21;527:110822. doi: 10.1016/j.jtbi.2021.110822. Epub 2021 Jun 29. J Theor Biol. 2021. PMID: 34214567
-
Non-enzymatic primer extension with strand displacement.Elife. 2019 Nov 8;8:e51888. doi: 10.7554/eLife.51888. Elife. 2019. PMID: 31702557 Free PMC article.
-
The evolution of strand preference in simulated RNA replicators with strand displacement: implications for the origin of transcription.Biol Direct. 2008 Aug 11;3:33. doi: 10.1186/1745-6150-3-33. Biol Direct. 2008. PMID: 18694481 Free PMC article.
-
Pathways of human cell post-replication repair.Carcinogenesis. 1989 Jan;10(1):1-11. doi: 10.1093/carcin/10.1.1. Carcinogenesis. 1989. PMID: 2642748 Review.
-
Transcription and replication of the influenza a virus genome.Acta Virol. 2000 Oct;44(5):273-82. Acta Virol. 2000. PMID: 11252672 Review.
Cited by
-
Peptide-based coacervates in therapeutic applications.Front Bioeng Biotechnol. 2023 Jan 4;10:1100365. doi: 10.3389/fbioe.2022.1100365. eCollection 2022. Front Bioeng Biotechnol. 2023. PMID: 36686257 Free PMC article. Review.
-
Simple peptides derived from the ribosomal core potentiate RNA polymerase ribozyme function.Nat Chem. 2017 Apr;9(4):325-332. doi: 10.1038/nchem.2739. Epub 2017 Mar 6. Nat Chem. 2017. PMID: 28338682 Free PMC article.
-
From chemical metabolism to life: the origin of the genetic coding process.Beilstein J Org Chem. 2017 Jun 12;13:1119-1135. doi: 10.3762/bjoc.13.111. eCollection 2017. Beilstein J Org Chem. 2017. PMID: 28684991 Free PMC article. Review.
-
N-Carboxyanhydride-Mediated Fatty Acylation of Amino Acids and Peptides for Functionalization of Protocell Membranes.J Am Chem Soc. 2016 Dec 28;138(51):16669-16676. doi: 10.1021/jacs.6b08801. Epub 2016 Dec 13. J Am Chem Soc. 2016. PMID: 27959544 Free PMC article.
-
Emergence of a New Self-Replicator from a Dynamic Combinatorial Library Requires a Specific Pre-Existing Replicator.J Am Chem Soc. 2017 Oct 4;139(39):13612-13615. doi: 10.1021/jacs.7b07346. Epub 2017 Sep 22. J Am Chem Soc. 2017. PMID: 28910535 Free PMC article.
References
-
- Orgel LE. Some consequences of the RNA world hypothesis. Orig Life Evol Biosph. 2003;33:211–218. - PubMed
-
- Gilbert W. Origin of life: the RNA world. Nature. 1986;319:618.
-
- Powner MW, Gerland B, Sutherland JD. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature. 2009;459:239–242. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
