

Review
doi: 10.1038/nrc.2016.84.
Oxygen availability and metabolic adaptations
Affiliations
- PMID: 27658636
- PMCID: PMC5546320
- DOI: 10.1038/nrc.2016.84
Item in Clipboard
Review
Oxygen availability and metabolic adaptations
Nat Rev Cancer.
.
Abstract
Oxygen availability, along with the abundance of nutrients (such as glucose, glutamine, lipids and albumin), fluctuates significantly during tumour evolution and the recruitment of blood vessels, leukocytes and reactive fibroblasts to complex tumour microenvironments. As such, hypoxia and concomitant nutrient scarcity affect large gene expression programmes, signalling pathways, diverse metabolic reactions and various stress responses. This Review summarizes our current understanding of how these adaptations are integrated in hypoxic tumour cells and their role in disease progression.
Figures

Left: Under normoxic conditions, HIF-1α and HIF-2α are degraded. The prolyl hydroxylase (PHD also referred to as “EgLN”) family of enzymes hydroxylate proline residues on the HIFα subunits, which are recognized by the pVHL subunit of an E3 ubiquitin ligase complex. Of note, this reaction requires oxygen, 2-oxoglutarate, ascorbate, and iron as cofactors. FIH hydroxylates an asparagine residue, inhibiting HIFα recruitment of p300/CBP cofactors. Of note, the precise contributions of ascorbate and iron to normoxic PHD2 enzymology is still under active investigation (see text). Right: Under hypoxic conditions, HIFα’s are not hydroxylated and instead translocate to the nucleus where they bind to their constitutively expressed partner ARNT. p300/CBP serve as transcriptional cofactors. HIFα/ARNT recognizes hypoxic response elements (HREs) throughout the genome and promote the transcription of 100s of genes involved in cellular adaptations to hypoxic stress. While HIF-1α and HIF-2α recognize the same HRE, they clearly promote the expression of mostly non-overlapping genes in certain contexts.

Lipids (e.g. fatty acids, cholesterol, phospholipids, and triglycerides) can be synthesized de novo from glucose or glutamine or taken up by cell surface transporters. Decreased oxygen levels promote glucose import, glycolysis, glutamine uptake, and glutaminolysis. Hypoxic cells also exhibit increased rates of reductive carboxylation. Moreover, hypoxia increases the expression of Fructose-1,6-bisphosphatase (FBP1). The tumor microenvironment can decrease stearoyl-CoA desaturase-1 (SCD1) desaturase activity. Thus, unsaturated lipids in particular are affected by changes in oxygen availability. Hypoxia results in increased lipid uptake, which counteracts the affects of SCD1 inhibition. Abbreviations: PKM1/2: pyruvate kinase M1/2; FASN: fatty acid synthase; HMGCR: HMG Co-A reductase; ACL: ATP citrate lyase; ACC: acetyl-CoA carboxylase.

Key activators (green) and inhibitors (red) of the mTORC1/2 pathway are shown. A diverse array of extracellular inputs, such as insulin, growth factors, and amino acids can promote mTOR activity. These signals lead to the inactivation of the TSC1/2 complex, an upstream inhibitor of mTORC1. They can also activate mTORC2 through PIP3 and AKT. For mTORC1, the main outputs are increased ribosomal biogenesis, CAP-dependent mRNA translation, and a series of anabolic pathways, such as purine, pyrimidine, and lipid synthesis. mTORC2 is associated with pro-survival activity, as well as modification of the cytoskeleton and other metabolic adaptations. Hypoxia can inhibit mTORC1 through REDD1 or REDD2, which are HIF-1α target genes. Additionally, hypoxia leads to energy deficiency (elevated AMP/ATP ratios), which can inhibit mTORC1 through AMPK. Interestingly, HIF-2α enhances mTORC2 activity by promoting amino acid import through the transporter SLC7A5. It should be noted that REDD1 can be phosphorylated and activated ahead of increased abundance downstream of HIF-1α, so it in and of itself is an “O2 sensor”.

Beyond the PHDs and FIH, 60–70 2-OG dependent dioxygenases have either been biochemically confirmed or predicted based on sequencing the human genome. Their relative KmO2 for molecular oxygen and dependence on Fe2+ indicate that their activities could increase or decrease, based on pO2 levels and concentrations of succinate, fumarate, and 2(L)-HG accumulation in the tumor microenvironment. As shown, they a have variety of enzymatic functions in cells which impact the extracellular matrix, methylation status of histones and nucleic acids, and components of the translation machinery, along with pivotal roles in maintaining oxygen homeostasis by hydroxylating HIFα subunits.
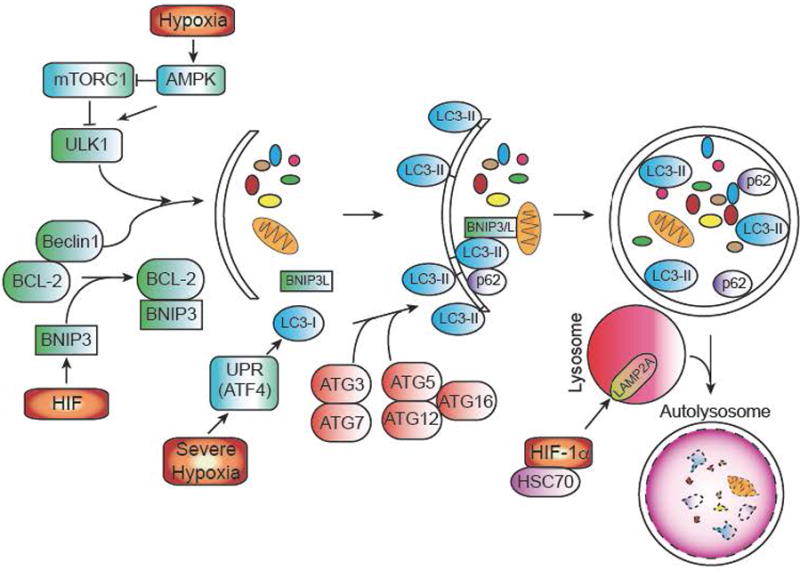
Autophagy (or macroautophagy) is a catabolic process in which double membrane vesicles called autophagosomes sequester and degrade organelles, proteins, and other cytoplasmic constituents in response to cellular stress. The autophagic pathway consists of several steps, including nucleation and sequestration of a phagophore, formation of an autophagosome, and its fusion with a lysosome to form an autolysosome. Initiation and nucleation of the autophagophore begins when several proteins, including ULK1 and Beclin-1, which are inhibited by mTORC1 activity and BCL-2 respectively. BNIP3, a hypoxically induced protein, displaces Beclin 1 from BCL-2. After nucleation, several ATG proteins are involved in the conjugation of phosphatidylethanolamine (PE) to LC3, to form LC3-II. LC3-II is necessary for autophagic membrane expansion, recognition of autophagic cargo, and the fusion of lysosomes to autophagosomes. Another hypoxically induced protein, BNIP3L, is involved in sequestering mitochondria to the autophagosome. p62 functions as an autophagy cargo receptor, which is ultimately degraded when autolysosome forms. Severe hypoxia (<.01% O2) can also activate autophagy in a HIF-independent mechanism through the UPR. Once the autophagosome has formed, it fuses with a lysosome to generate an autolysosome, leading to the degradation of the products inside. Autophagy can also modulate HIF-1α levels as well. Chaperone-mediated autophagy proteins HSC70 and LAMP2A traffic HIF-1α to lysosomes, where it is subsequently degraded.
References
-
- Lee KE, Simon MC. Snapshot: Hypoxia-Inducible Factors. Cell. 2015 in press. - PubMed
-
- Brizel DM, et al. Tumor oxygenation predicts for the likelihood of distant metastases in human soft tissue sarcoma. Cancer Res. 1996;56:941–943. - PubMed
-
- Hockel M, et al. Intratumoral pO2 predicts survival in advanced cancer of the uterine cervix. Radiother Oncol. 1993;26:45–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
