

Establishment and functions of DNA methylation in the germline
- PMID: 27659720
- PMCID: PMC5066131
- DOI: 10.2217/epi-2016-0056
Establishment and functions of DNA methylation in the germline
Abstract
Epigenetic modifications established during gametogenesis regulate transcription and other nuclear processes in gametes, but also have influences in the zygote, embryo and postnatal life. This is best understood for DNA methylation which, established at discrete regions of the oocyte and sperm genomes, governs genomic imprinting. In this review, we describe how imprinting has informed our understanding of de novo DNA methylation mechanisms, highlight how recent genome-wide profiling studies have provided unprecedented insights into establishment of the sperm and oocyte methylomes and consider the fate and function of gametic methylation and other epigenetic modifications after fertilization.
Keywords: DNA methylation; embryo; imprinting; oogenesis; spermatogenesis; transgenerational inheritance.
Conflict of interest statement
Financial & competing interests disclosure KR Stewart was supported by a studentship from the Cambridge Overseas Trusts. L Veselovska was supported by a Babraham Institute: Cambridge University studentship. Work in G Kelsey's laboratory is funded by the Biotechnology and Biological Sciences Research Council and the Medical Research Council. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed. No writing assistance was utilized in the production of this manuscript.
Figures
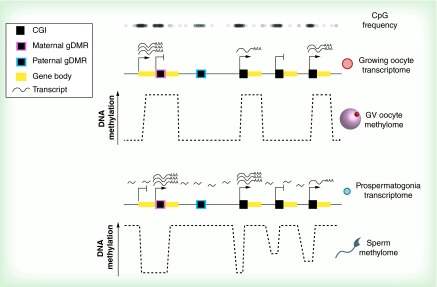
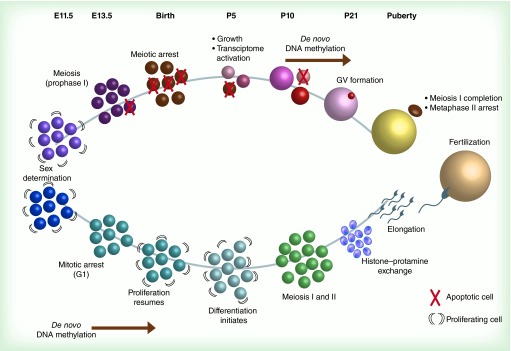
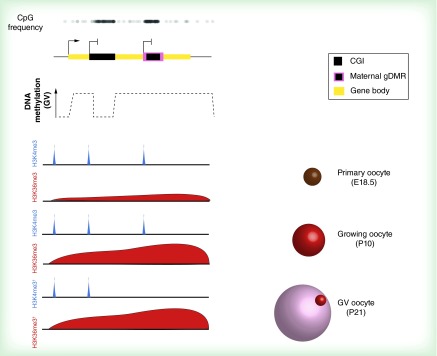
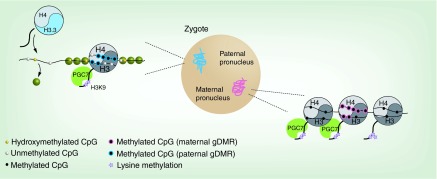
References
-
- Jones PA. Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012;13(7):484–492. - PubMed
-
- Chen T, Li E. Structure and function of eukaryotic DNA methyltransferases. Curr. Top. Dev. Biol. 2004;60:55–89. - PubMed
-
- Hitt MM, Wu TL, Cohen G, Linn S. De novo and maintenance DNA methylation by a mouse plasmacytoma cell DNA methyltransferase. J. Biol. Chem. 1988;263(9):4392–4399. - PubMed
-
- Pradhan S, Bacolla A, Wells RD, Roberts RJ. Recombinant human DNA (cytosine-5) methyltransferase: I. Expression, purification, and comparison of de novo and maintenance methylation. J. Biol. Chem. 1999;274(46):33002–33010. - PubMed
-
- Arita K, Ariyoshi M, Tochio H, Nakamura Y, Shirakawa M. Recognition of hemi-methylated DNA by the SRA protein UHRF1 by a base-flipping mechanism. Nature. 2008;455(7214):818–821. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources