

Editing DNA Methylation in the Mammalian Genome
- PMID: 27662091
- PMCID: PMC5062609
- DOI: 10.1016/j.cell.2016.08.056
Editing DNA Methylation in the Mammalian Genome
Abstract
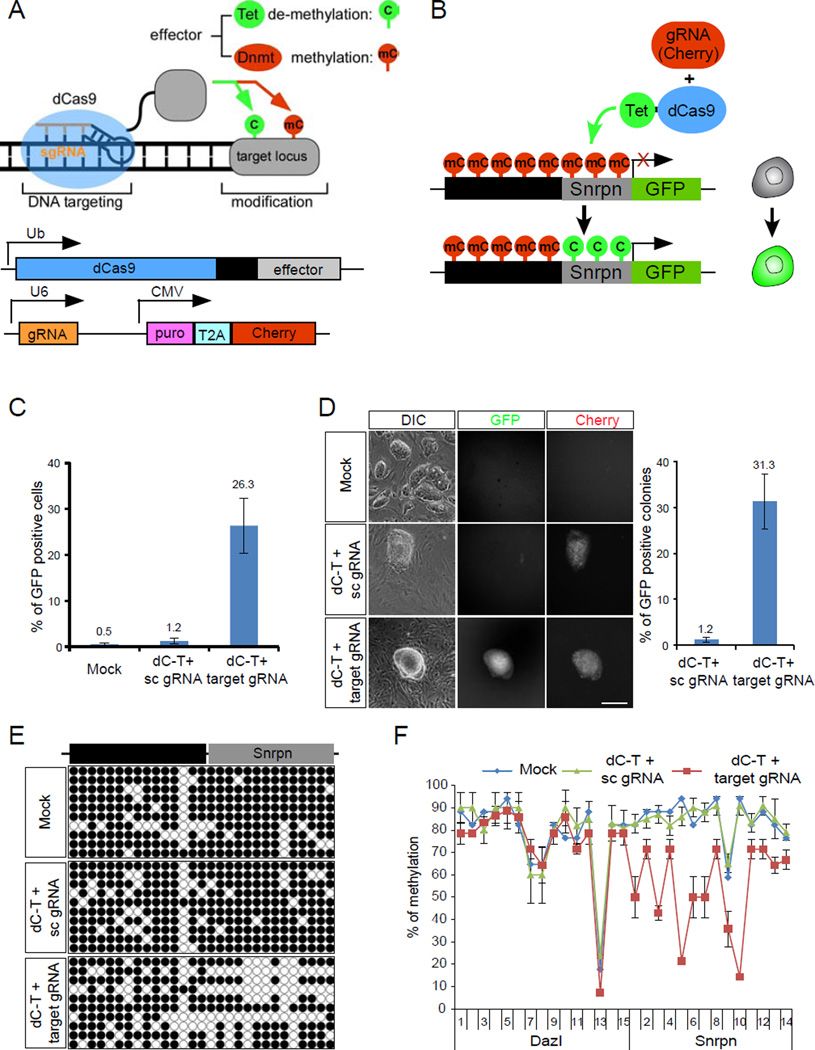
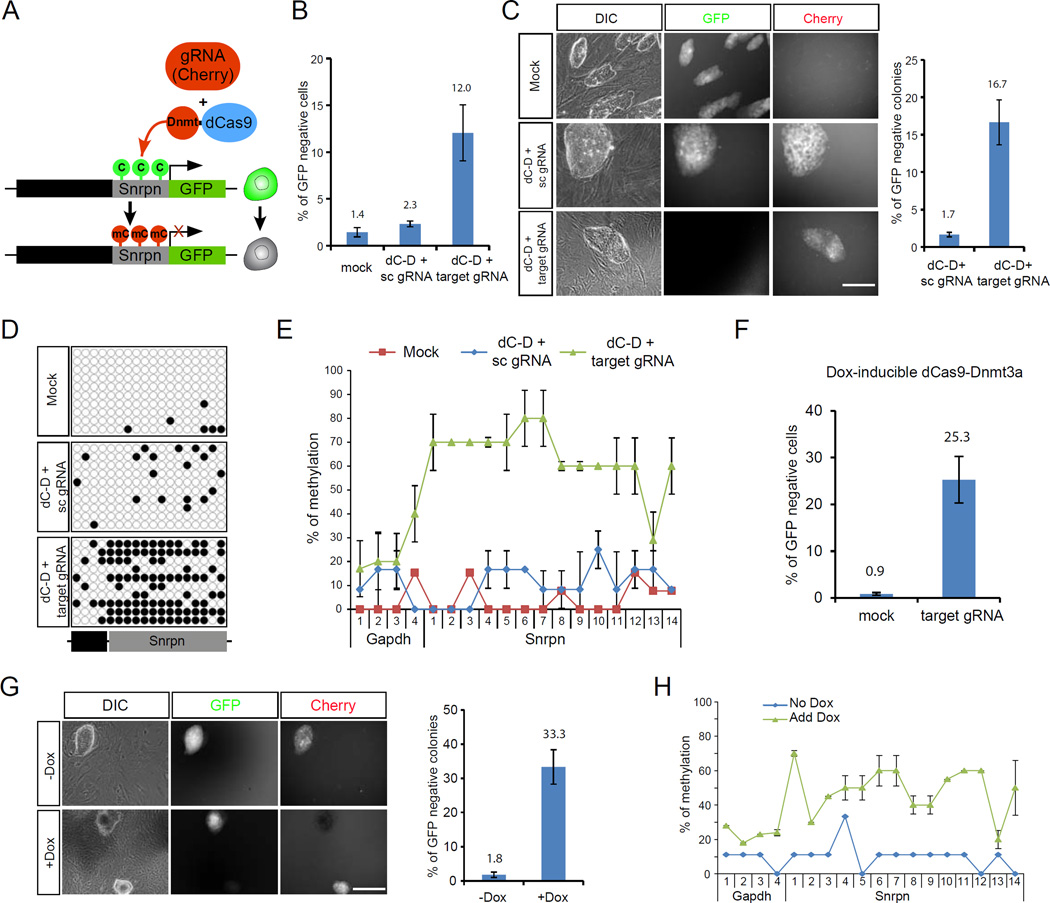
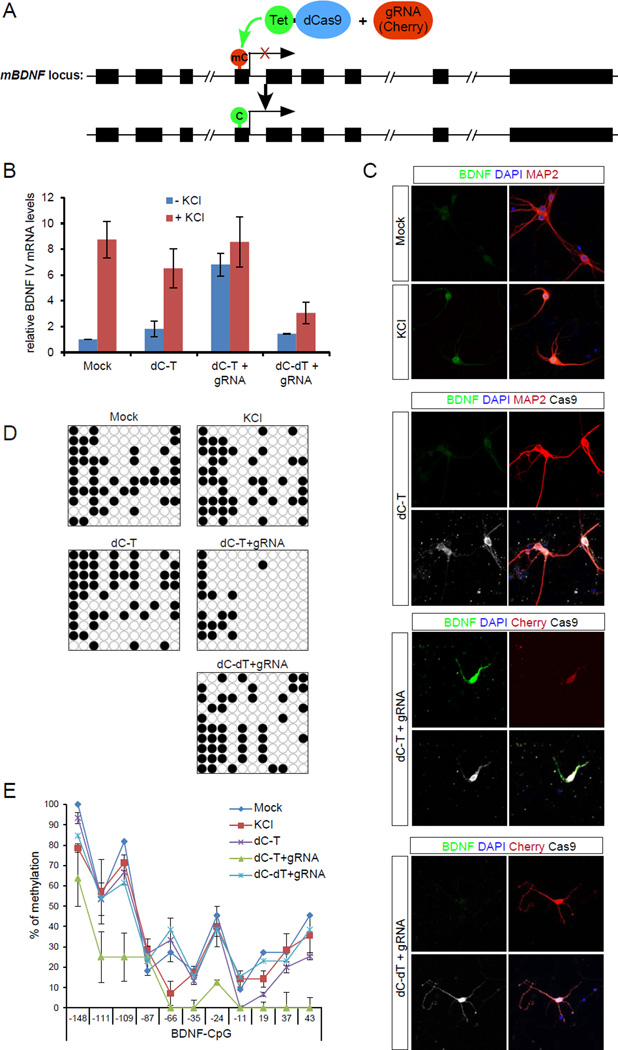
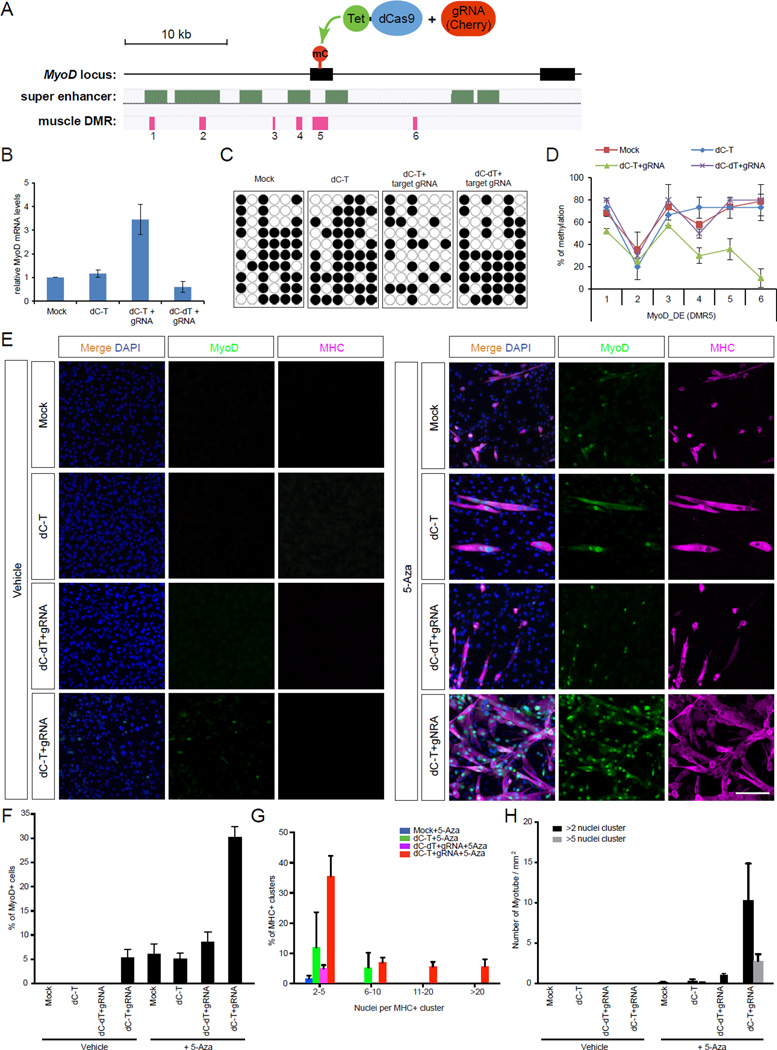
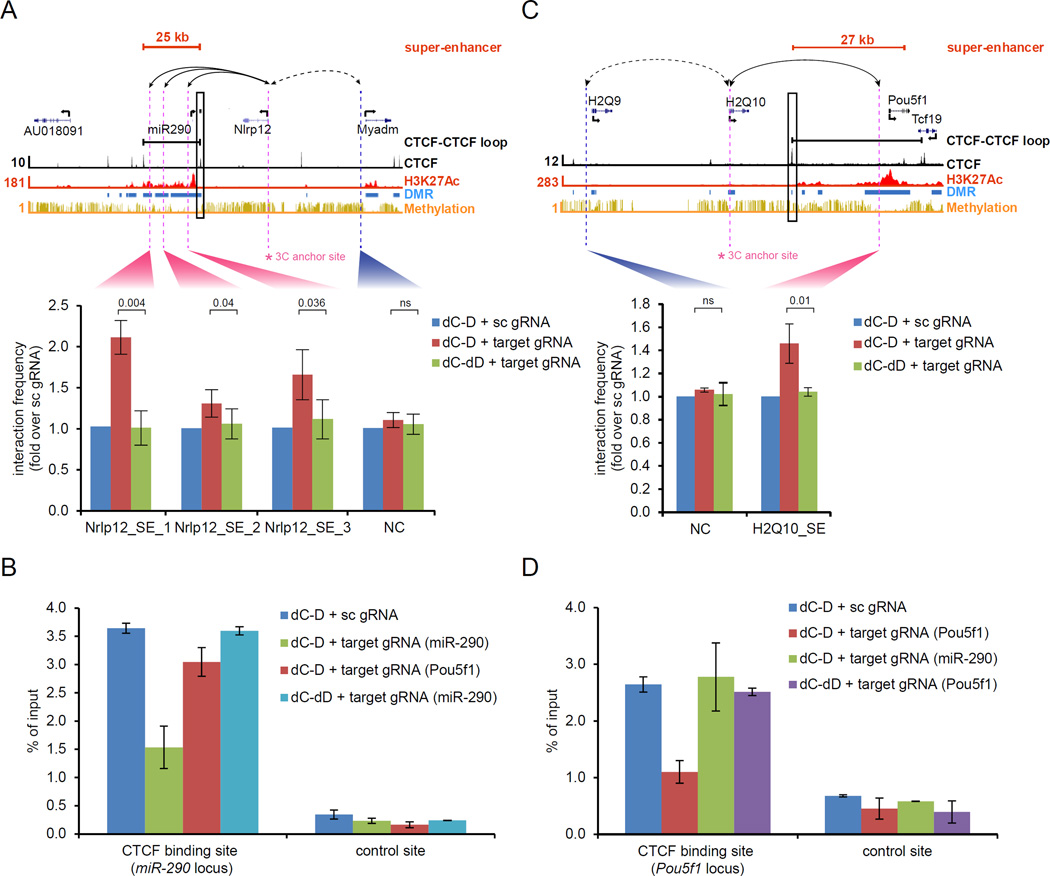
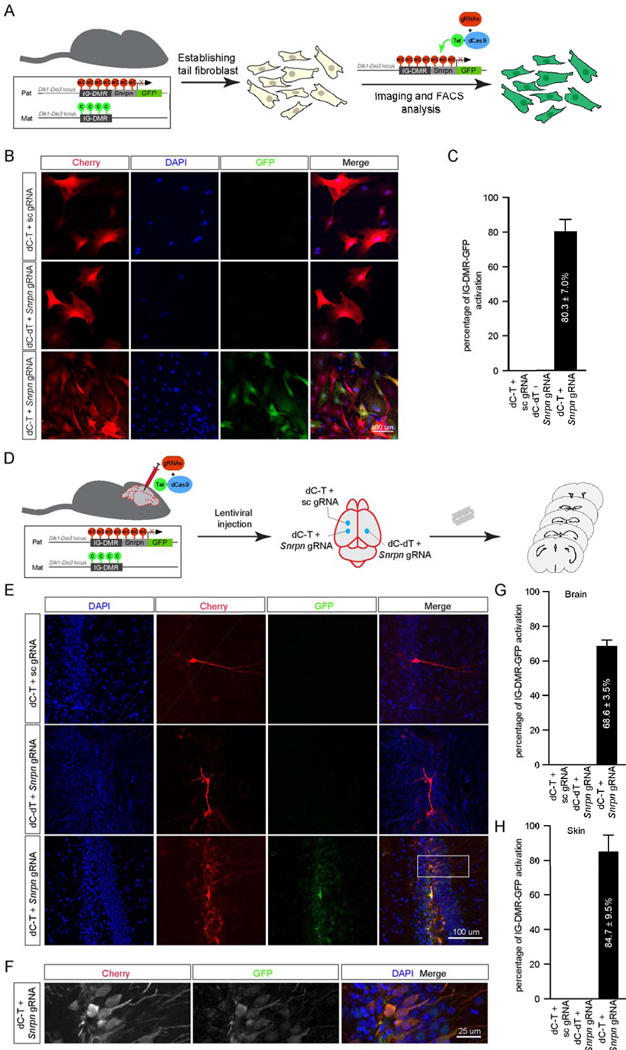
Mammalian DNA methylation is a critical epigenetic mechanism orchestrating gene expression networks in many biological processes. However, investigation of the functions of specific methylation events remains challenging. Here, we demonstrate that fusion of Tet1 or Dnmt3a with a catalytically inactive Cas9 (dCas9) enables targeted DNA methylation editing. Targeting of the dCas9-Tet1 or -Dnmt3a fusion protein to methylated or unmethylated promoter sequences caused activation or silencing, respectively, of an endogenous reporter. Targeted demethylation of the BDNF promoter IV or the MyoD distal enhancer by dCas9-Tet1 induced BDNF expression in post-mitotic neurons or activated MyoD facilitating reprogramming of fibroblasts into myoblasts, respectively. Targeted de novo methylation of a CTCF loop anchor site by dCas9-Dnmt3a blocked CTCF binding and interfered with DNA looping, causing altered gene expression in the neighboring loop. Finally, we show that these tools can edit DNA methylation in mice, demonstrating their wide utility for functional studies of epigenetic regulation.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures

References
-
- Bell AC, Felsenfeld G. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature. 2000;405:482–485. - PubMed
-
- Bird A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002;16:6–21. - PubMed
-
- Brunk BP, Goldhamer DJ, Emerson CP., Jr Regulated demethylation of the myoD distal enhancer during skeletal myogenesis. Dev Biol. 1996;177:490–503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
