

Overlapping Role of Respiratory Supercomplex Factor Rcf2 and Its N-terminal Homolog Rcf3 in Saccharomyces cerevisiae
- PMID: 27662906
- PMCID: PMC5095429
- DOI: 10.1074/jbc.M116.734665
Overlapping Role of Respiratory Supercomplex Factor Rcf2 and Its N-terminal Homolog Rcf3 in Saccharomyces cerevisiae
Abstract
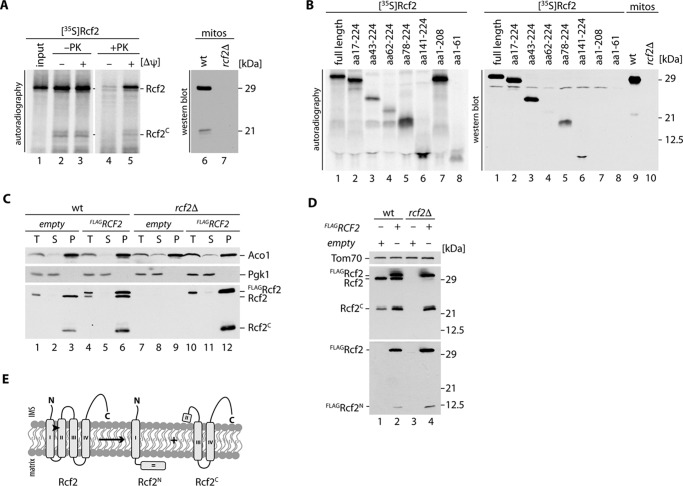
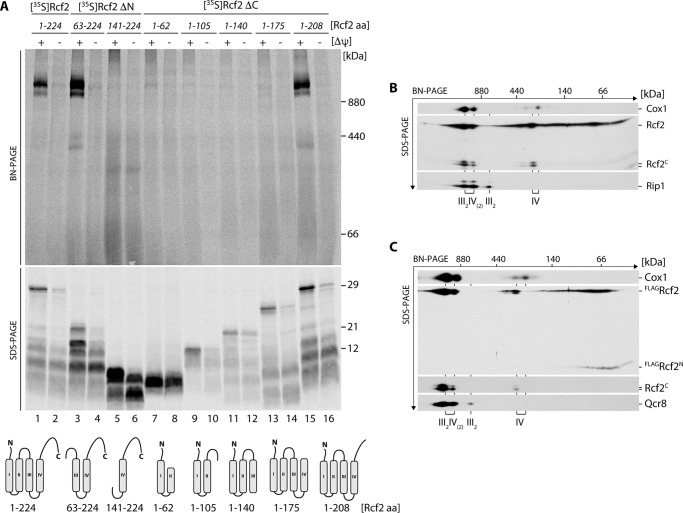
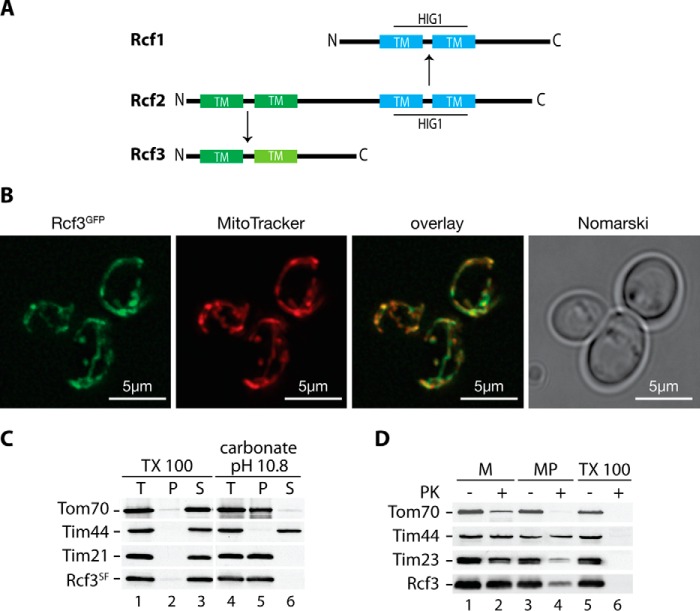
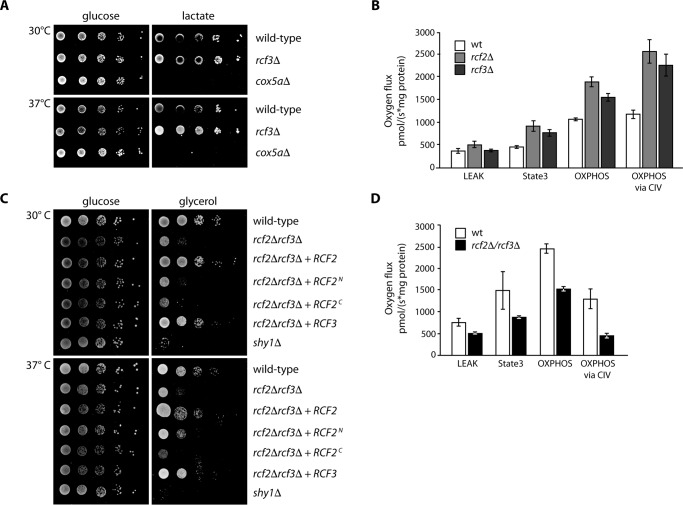
The mitochondrial electron transport chain consists of individual protein complexes arranged into large macromolecular structures, termed respiratory chain supercomplexes or respirasomes. In the yeast Saccharomyces cerevisiae, respiratory chain supercomplexes form by association of the bc1 complex with the cytochrome c oxidase. Formation and maintenance of these assemblies are promoted by specific respiratory supercomplex factors, the Rcf proteins. For these proteins a regulatory function in bridging the electron transfer within supercomplexes has been proposed. Here we report on the maturation of Rcf2 into an N- and C-terminal peptide. We show that the previously uncharacterized Rcf3 (YBR255c-A) is a homolog of the N-terminal Rcf2 peptide, whereas Rcf1 is homologous to the C-terminal portion. Both Rcf3 and the C-terminal fragment of Rcf2 associate with monomeric cytochrome c oxidase and respiratory chain supercomplexes. A lack of Rcf2 and Rcf3 increases oxygen flux through the respiratory chain by up-regulation of the cytochrome c oxidase activity. A double gene deletion of RCF2 and RCF3 affects cellular survival under non-fermentable growth conditions, suggesting an overlapping role for both proteins in the regulation of the OXPHOS activity. Furthermore, our data suggest an association of all three Rcf proteins with the bc1 complex in the absence of a functional cytochrome c oxidase and identify a supercomplex independent interaction network of the Rcf proteins.
Keywords: cytochrome c oxidase (complex IV); membrane protein; mitochondria; protein assembly; respiratory chain.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Rcf proteins and their differential specificity for respiratory chain complexes: A unique role for Rcf2 on oxygen sensitive supercomplexes?Biochim Biophys Acta Mol Cell Res. 2021 Nov;1868(12):119133. doi: 10.1016/j.bbamcr.2021.119133. Epub 2021 Aug 25. Biochim Biophys Acta Mol Cell Res. 2021. PMID: 34450214
-
The yeast mitochondrial proteins Rcf1 and Rcf2 support the enzymology of the cytochrome c oxidase complex and generation of the proton motive force.J Biol Chem. 2019 Mar 29;294(13):4867-4877. doi: 10.1074/jbc.RA118.006888. Epub 2019 Jan 25. J Biol Chem. 2019. PMID: 30683696 Free PMC article.
-
Rcf1 mediates cytochrome oxidase assembly and respirasome formation, revealing heterogeneity of the enzyme complex.Cell Metab. 2012 Mar 7;15(3):336-47. doi: 10.1016/j.cmet.2012.01.016. Epub 2012 Feb 16. Cell Metab. 2012. PMID: 22342701
-
HIGD-Driven Regulation of Cytochrome c Oxidase Biogenesis and Function.Cells. 2020 Dec 6;9(12):2620. doi: 10.3390/cells9122620. Cells. 2020. PMID: 33291261 Free PMC article. Review.
-
Mitochondrial respiratory supercomplexes of the yeast Saccharomyces cerevisiae.IUBMB Life. 2024 Aug;76(8):485-504. doi: 10.1002/iub.2817. Epub 2024 Mar 26. IUBMB Life. 2024. PMID: 38529880 Review.
Cited by
-
EARLY NODULIN93 acts via cytochrome c oxidase to alter respiratory ATP production and root growth in plants.Plant Cell. 2024 Nov 2;36(11):4716-4731. doi: 10.1093/plcell/koae242. Plant Cell. 2024. PMID: 39179507 Free PMC article.
-
Structure and function of the S. pombe III-IV-cyt c supercomplex.Proc Natl Acad Sci U S A. 2023 Nov 14;120(46):e2307697120. doi: 10.1073/pnas.2307697120. Epub 2023 Nov 8. Proc Natl Acad Sci U S A. 2023. PMID: 37939086 Free PMC article.
-
The Cox1 C-terminal domain is a central regulator of cytochrome c oxidase biogenesis in yeast mitochondria.J Biol Chem. 2017 Jun 30;292(26):10912-10925. doi: 10.1074/jbc.M116.773077. Epub 2017 May 10. J Biol Chem. 2017. PMID: 28490636 Free PMC article.
-
Keep Calm and Survive: Adaptation Strategies to Energy Crisis in Fruit Trees under Root Hypoxia.Plants (Basel). 2020 Aug 27;9(9):1108. doi: 10.3390/plants9091108. Plants (Basel). 2020. PMID: 32867316 Free PMC article. Review.
-
Structure and Mechanism of Respiratory III-IV Supercomplexes in Bioenergetic Membranes.Chem Rev. 2021 Aug 11;121(15):9644-9673. doi: 10.1021/acs.chemrev.1c00140. Epub 2021 Jun 29. Chem Rev. 2021. PMID: 34184881 Free PMC article. Review.
References
-
- Grandier-Vazeille X., Bathany K., Chaignepain S., Camougrand N., Manon S., and Schmitter J. M. (2001) Yeast mitochondrial dehydrogenases are associated in a supramolecular complex. Biochemistry 40, 9758–9769 - PubMed
-
- Zara V., Conte L., and Trumpower B. L. (2009) Biogenesis of the yeast cytochrome bc1 complex. Biochim. Biophys. Acta. 1793, 89–96 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
