

The RNA-binding protein vigilin regulates VLDL secretion through modulation of Apob mRNA translation
- PMID: 27665711
- PMCID: PMC5052685
- DOI: 10.1038/ncomms12848
The RNA-binding protein vigilin regulates VLDL secretion through modulation of Apob mRNA translation
Abstract
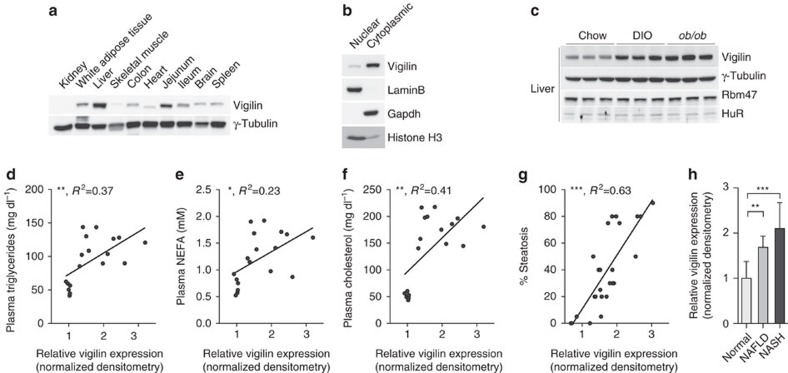
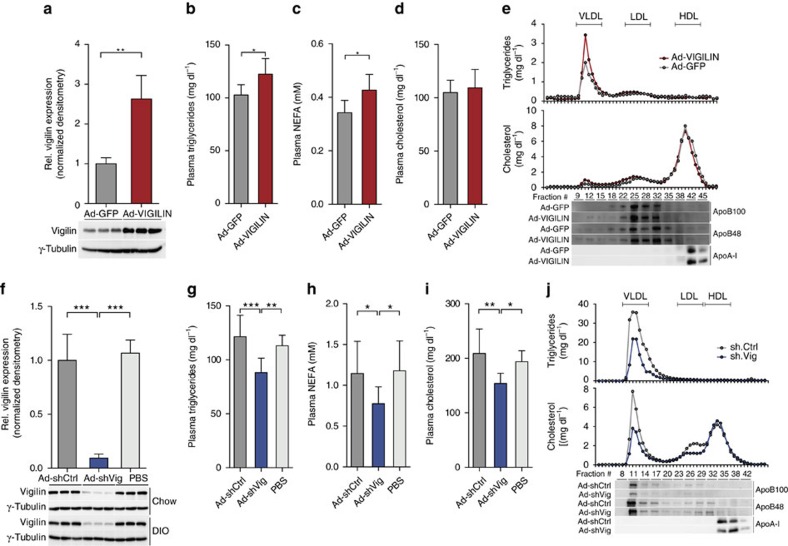
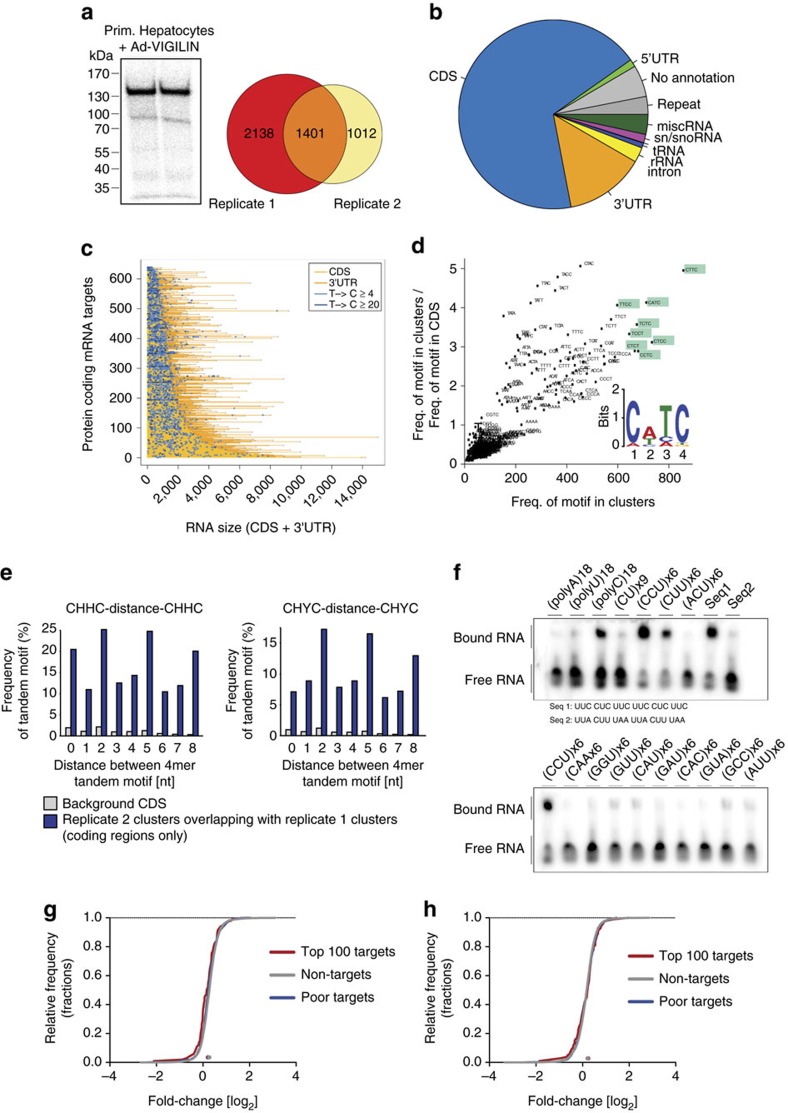
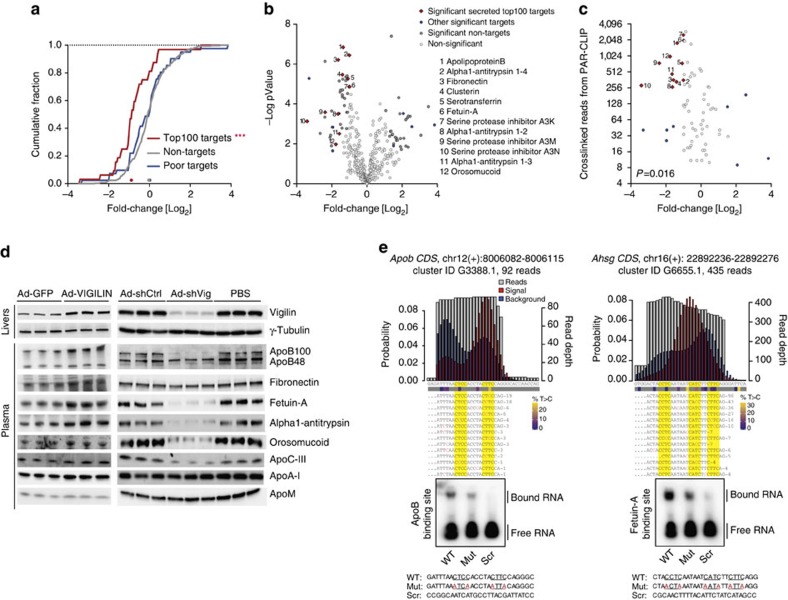
The liver is essential for the synthesis of plasma proteins and integration of lipid metabolism. While the role of transcriptional networks in these processes is increasingly understood, less is known about post-transcriptional control of gene expression by RNA-binding proteins (RBPs). Here, we show that the RBP vigilin is upregulated in livers of obese mice and in patients with fatty liver disease. By using in vivo, biochemical and genomic approaches, we demonstrate that vigilin controls very-low-density lipoprotein (VLDL) secretion through the modulation of apolipoproteinB/Apob mRNA translation. Crosslinking studies reveal that vigilin binds to CU-rich regions in the mRNA coding sequence of Apob and other proatherogenic secreted proteins, including apolipoproteinC-III/Apoc3 and fibronectin/Fn1. Consequently, hepatic vigilin knockdown decreases VLDL/low-density lipoprotein (LDL) levels and formation of atherosclerotic plaques in Ldlr-/- mice. These studies uncover a role for vigilin as a key regulator of hepatic Apob translation and demonstrate the therapeutic potential of inhibiting vigilin for cardiovascular diseases.
Conflict of interest statement
M.S. and T.T. are members of the Scientific Advisory Board and M.M. and K.C. are employees of Alnylam Pharmaceuticals. The other authors declare no competing financial interests.
Figures
Similar articles
-
microRNA-185 modulates low density lipoprotein receptor expression as a key posttranscriptional regulator.Atherosclerosis. 2015 Dec;243(2):523-32. doi: 10.1016/j.atherosclerosis.2015.10.026. Epub 2015 Oct 23. Atherosclerosis. 2015. PMID: 26523989
-
The magnitude of decrease in hepatic very low density lipoprotein apolipoprotein B secretion is determined by the extent of 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibition in miniature pigs.Endocrinology. 1999 Nov;140(11):5293-302. doi: 10.1210/endo.140.11.7150. Endocrinology. 1999. PMID: 10537160
-
RNAi-mediated depletion of the 15 KH domain protein, vigilin, induces death of dividing and non-dividing human cells but does not initially inhibit protein synthesis.Nucleic Acids Res. 2003 Oct 1;31(19):5644-53. doi: 10.1093/nar/gkg768. Nucleic Acids Res. 2003. PMID: 14500828 Free PMC article.
-
APOLIPOPROTEIN B: mRNA editing, lipoprotein assembly, and presecretory degradation.Annu Rev Nutr. 2000;20:169-93. doi: 10.1146/annurev.nutr.20.1.169. Annu Rev Nutr. 2000. PMID: 10940331 Review.
-
A jack of all trades: the RNA-binding protein vigilin.Wiley Interdiscip Rev RNA. 2017 Nov;8(6). doi: 10.1002/wrna.1448. Epub 2017 Oct 4. Wiley Interdiscip Rev RNA. 2017. PMID: 28975734 Review.
Cited by
-
Heparin-binding EGF-like growth factor (HB-EGF) antisense oligonucleotide protected against hyperlipidemia-associated atherosclerosis.Nutr Metab Cardiovasc Dis. 2019 Mar;29(3):306-315. doi: 10.1016/j.numecd.2018.12.006. Epub 2019 Jan 9. Nutr Metab Cardiovasc Dis. 2019. PMID: 30738642 Free PMC article.
-
In vivo PAR-CLIP (viP-CLIP) of liver TIAL1 unveils targets regulating cholesterol synthesis and secretion.Nat Commun. 2023 Jun 9;14(1):3386. doi: 10.1038/s41467-023-39135-8. Nat Commun. 2023. PMID: 37296170 Free PMC article.
-
"SERBP1 (Hero45) is a Novel Link with Ischemic Heart Disease Risk: Associations with Coronary Arteries Occlusion, Blood Coagulation and Lipid Profile".Cell Biochem Biophys. 2025 Apr 3. doi: 10.1007/s12013-025-01736-z. Online ahead of print. Cell Biochem Biophys. 2025. PMID: 40175693
-
Alternative splicing and related RNA binding proteins in human health and disease.Signal Transduct Target Ther. 2024 Feb 2;9(1):26. doi: 10.1038/s41392-024-01734-2. Signal Transduct Target Ther. 2024. PMID: 38302461 Free PMC article. Review.
-
Identification of Vigilin as a Potential Ischemia Biomarker by Brain Slice-Based Systematic Evolution of Ligands by Exponential Enrichment.Anal Chem. 2019 May 21;91(10):6675-6681. doi: 10.1021/acs.analchem.9b00609. Epub 2019 May 1. Anal Chem. 2019. PMID: 30993971 Free PMC article.
References
-
- McKnight G. L. et al. Cloning and expression of a cellular high density lipoprotein-binding protein that is up-regulated by cholesterol loading of cells. J. Biol. Chem. 267, 12131–12141 (1992). - PubMed
-
- Schmidt C. et al. Complete cDNA sequence of chicken vigilin, a novel protein with amplified and evolutionary conserved domains. Eur. J. Biochem. 206, 625–634 (1992). - PubMed
-
- Burd C. G. & Dreyfuss G. Conserved structures and diversity of functions of RNA-binding proteins. Science 265, 615–621 (1994). - PubMed
-
- Dejgaard K. & Leffers H. Characterisation of the nucleic-acid-binding activity of KH domains. Different properties of different domains. Eur. J. Biochem. 241, 425–431 (1996). - PubMed
-
- Dodson R. E. & Shapiro D. J. Vigilin, a ubiquitous protein with 14 K homology domains, is the estrogen-inducible vitellogenin mRNA 3'-untranslated region-binding protein. J. Biol. Chem. 272, 12249–12252 (1997). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
