

Vimentin Intermediate Filaments Template Microtubule Networks to Enhance Persistence in Cell Polarity and Directed Migration
- PMID: 27667364
- PMCID: PMC5055390
- DOI: 10.1016/j.cels.2016.08.007
Vimentin Intermediate Filaments Template Microtubule Networks to Enhance Persistence in Cell Polarity and Directed Migration
Erratum in
-
Vimentin Intermediate Filaments Template Microtubule Networks to Enhance Persistence in Cell Polarity and Directed Migration.Cell Syst. 2016 Nov 23;3(5):500-501. doi: 10.1016/j.cels.2016.11.011. Cell Syst. 2016. PMID: 27883892 No abstract available.
Abstract
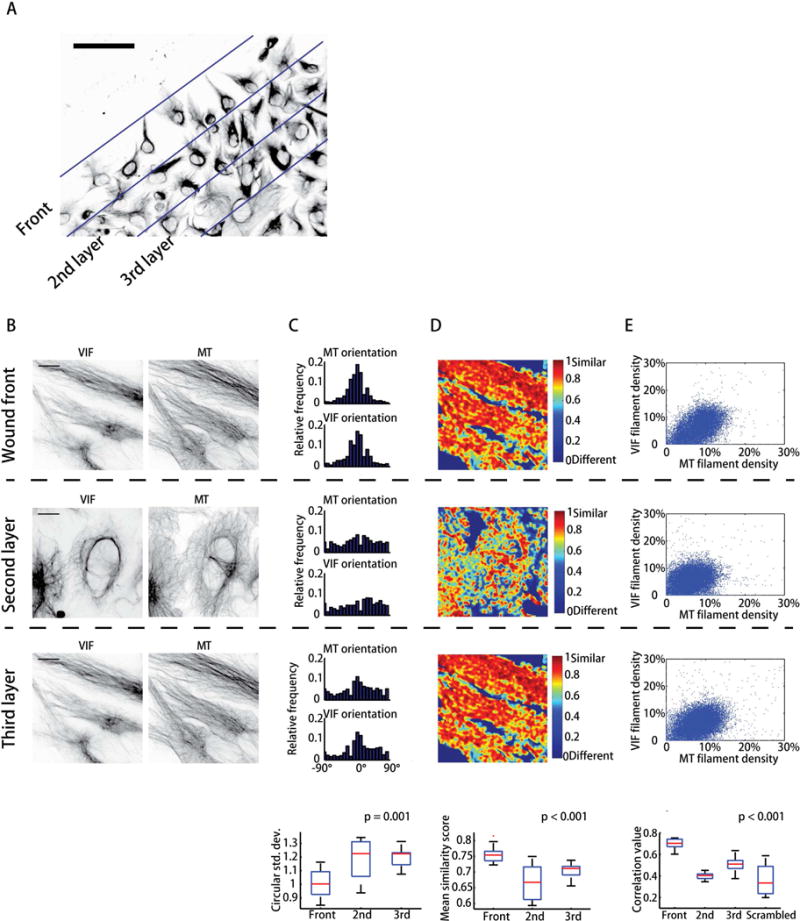
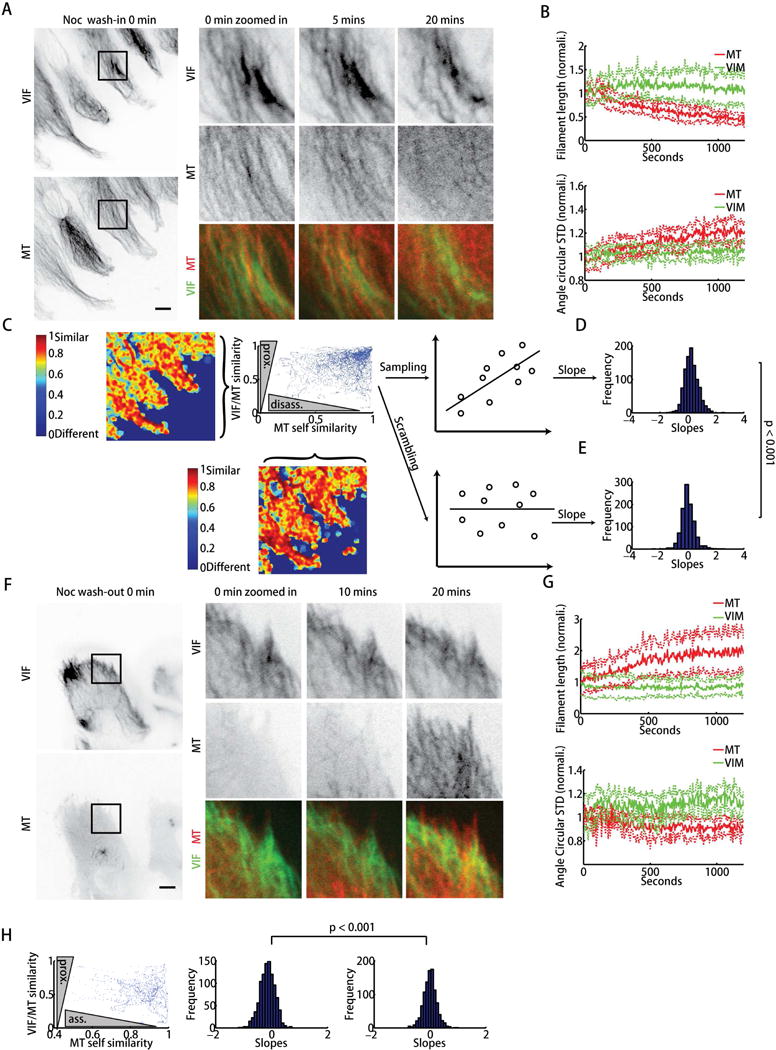
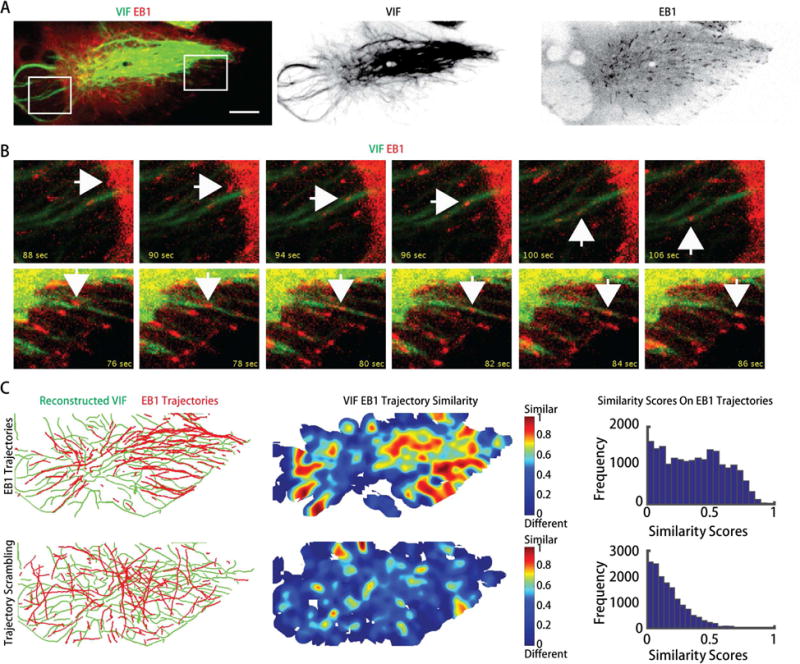
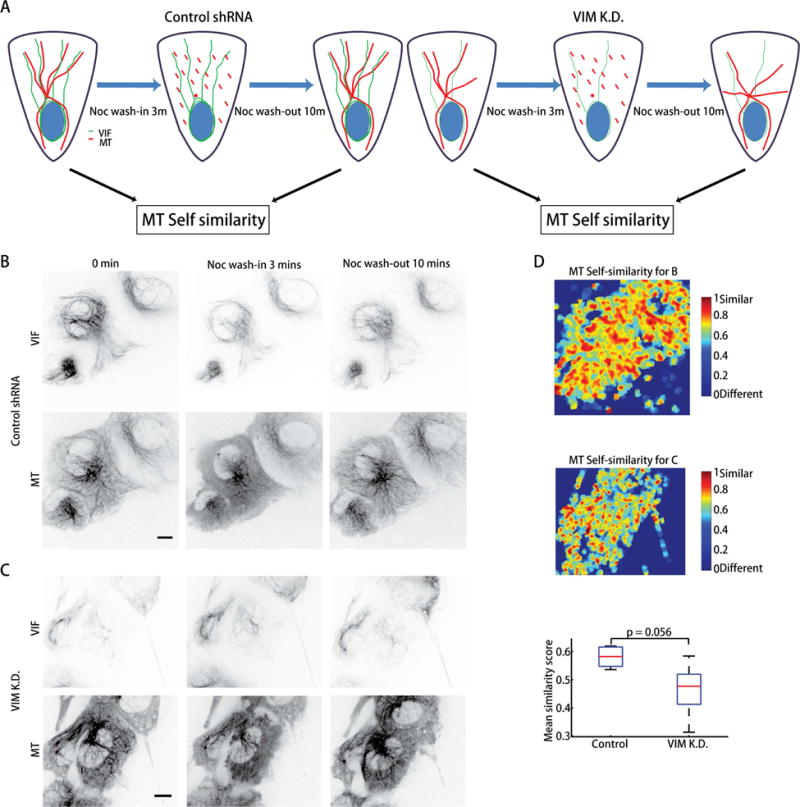
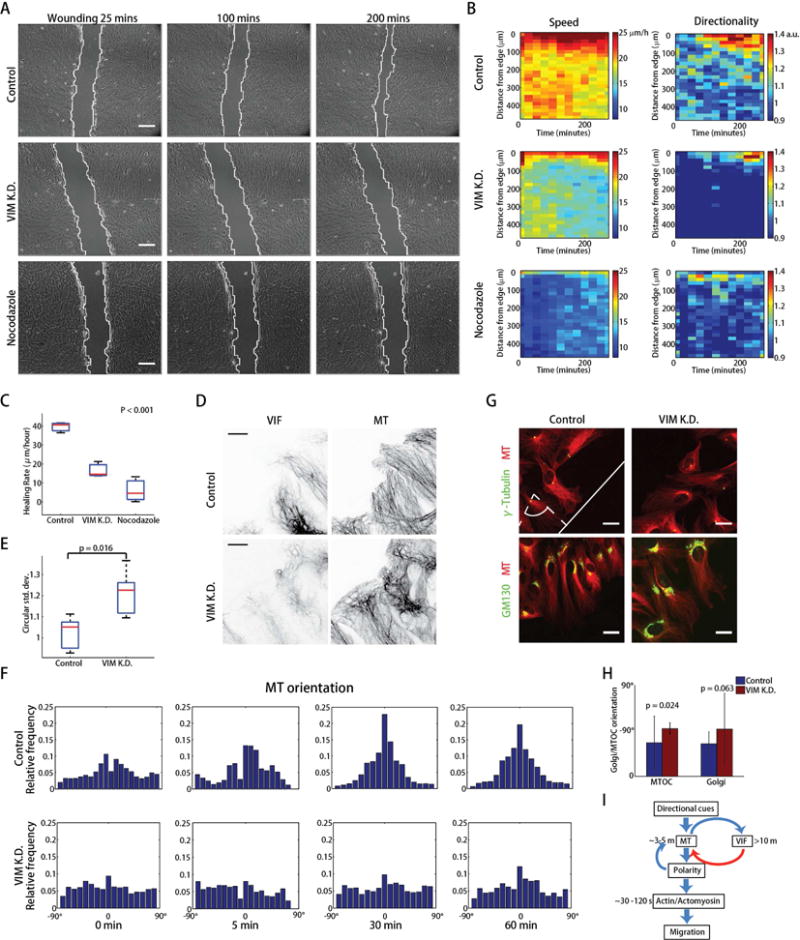
Increased expression of vimentin intermediate filaments (VIFs) enhances directed cell migration, but the mechanism behind VIFs' effect on motility is not understood. VIFs interact with microtubules, whose organization contributes to polarity maintenance in migrating cells. Here, we characterize the dynamic coordination of VIF and microtubule networks in wounded monolayers of retinal pigment epithelial cells. By genome editing, we fluorescently labeled endogenous vimentin and α-tubulin, and we developed computational image analysis to delineate architecture and interactions of the two networks. Our results show that VIFs assemble an ultrastructural copy of the previously polarized microtubule network. Because the VIF network is long-lived compared to the microtubule network, VIFs template future microtubule growth along previous microtubule tracks, thus providing a feedback mechanism that maintains cell polarity. VIF knockdown prevents cells from polarizing and migrating properly during wound healing. We suggest that VIFs' templating function establishes a memory in microtubule organization that enhances persistence in cell polarization in general and migration in particular.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures

References
-
- Chang L, Goldman RD. Intermediate filaments mediate cytoskeletal crosstalk. Nature reviews Molecular cell biology. 2004;5:601–613. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
