

Mutations in Human Accelerated Regions Disrupt Cognition and Social Behavior
- PMID: 27667684
- PMCID: PMC5063026
- DOI: 10.1016/j.cell.2016.08.071
Mutations in Human Accelerated Regions Disrupt Cognition and Social Behavior
Abstract
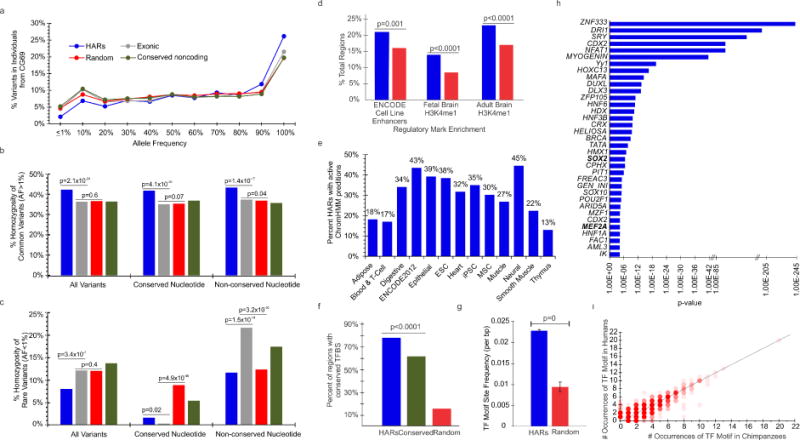
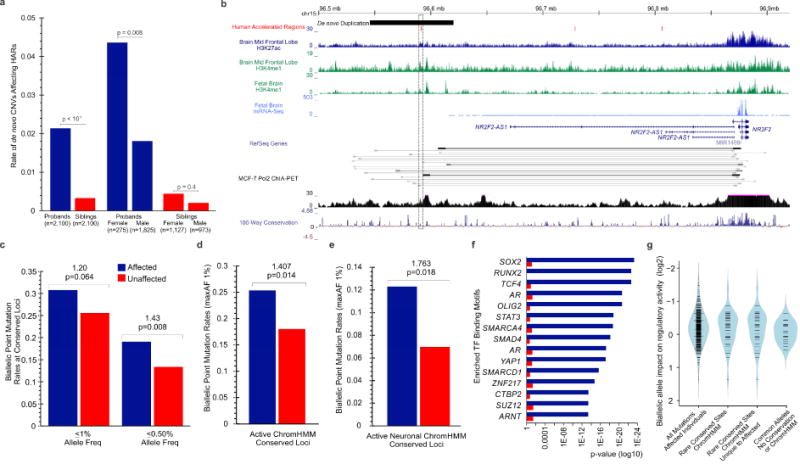
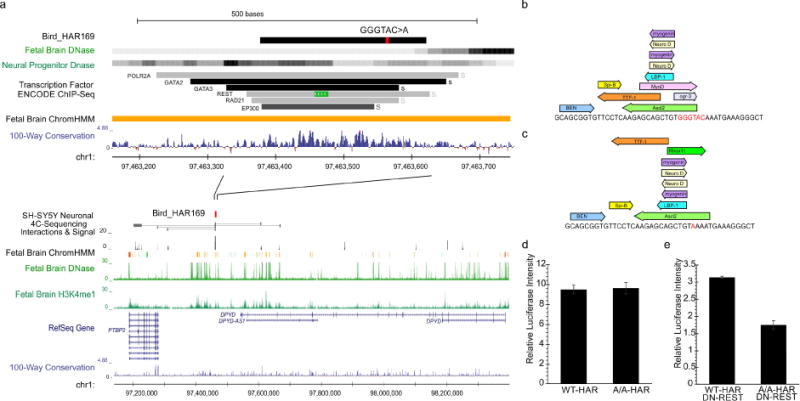
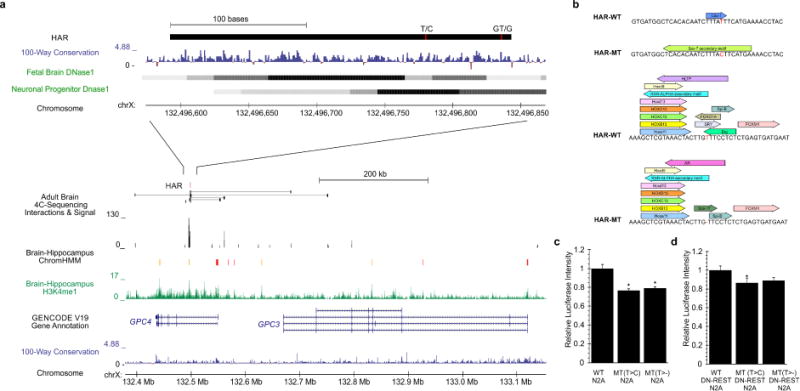
Comparative analyses have identified genomic regions potentially involved in human evolution but do not directly assess function. Human accelerated regions (HARs) represent conserved genomic loci with elevated divergence in humans. If some HARs regulate human-specific social and behavioral traits, then mutations would likely impact cognitive and social disorders. Strikingly, rare biallelic point mutations-identified by whole-genome and targeted "HAR-ome" sequencing-showed a significant excess in individuals with ASD whose parents share common ancestry compared to familial controls, suggesting a contribution in 5% of consanguineous ASD cases. Using chromatin interaction sequencing, massively parallel reporter assays (MPRA), and transgenic mice, we identified disease-linked, biallelic HAR mutations in active enhancers for CUX1, PTBP2, GPC4, CDKL5, and other genes implicated in neural function, ASD, or both. Our data provide genetic evidence that specific HARs are essential for normal development, consistent with suggestions that their evolutionary changes may have altered social and/or cognitive behavior. PAPERCLIP.
Keywords: ASD; Autism; Brain Evolution; HARs; Human Accelerated regions; noncoding.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures


Comment in
-
Darwin Comes to Clinic.Trends Genet. 2017 Jan;33(1):1-2. doi: 10.1016/j.tig.2016.11.005. Epub 2016 Nov 28. Trends Genet. 2017. PMID: 27908673
References
-
- Bienvenu T, Diebold B, Chelly J, Isidor B. Refining the phenotype associated with MEF2C point mutations. Neurogenetics. 2013;14:71–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
