

Segrosome Complex Formation during DNA Trafficking in Bacterial Cell Division
- PMID: 27668216
- PMCID: PMC5016525
- DOI: 10.3389/fmolb.2016.00051
Segrosome Complex Formation during DNA Trafficking in Bacterial Cell Division
Abstract
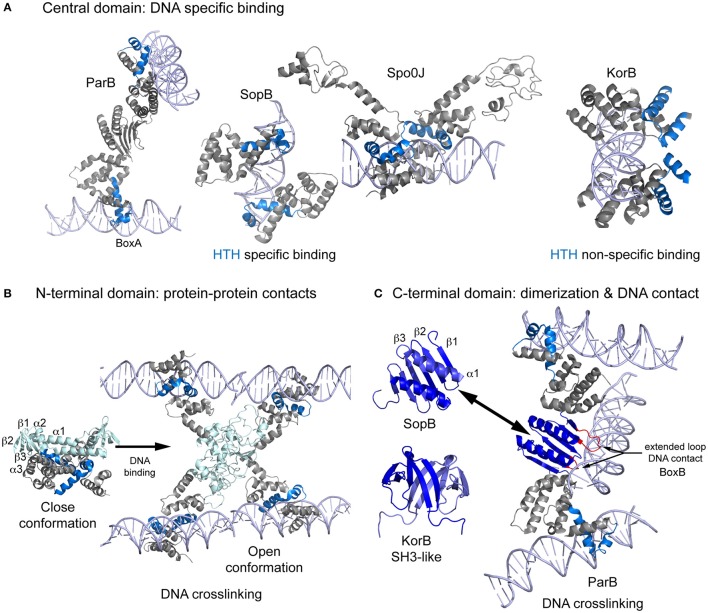
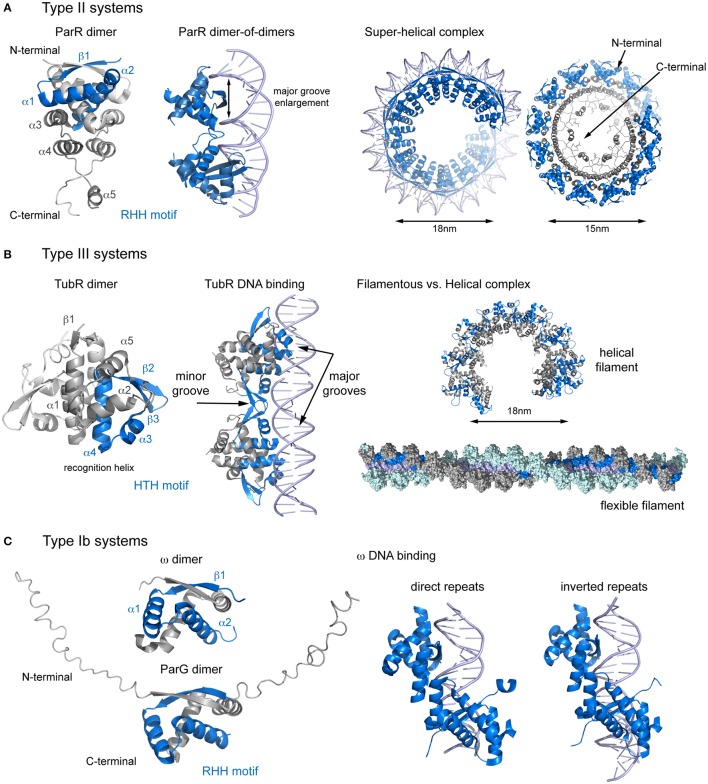
Bacterial extrachromosomal DNAs often contribute to virulence in pathogenic organisms or facilitate adaptation to particular environments. The transmission of genetic information from one generation to the next requires sufficient partitioning of DNA molecules to ensure that at least one copy reaches each side of the division plane and is inherited by the daughter cells. Segregation of the bacterial chromosome occurs during or after replication and probably involves a strategy in which several protein complexes participate to modify the folding pattern and distribution first of the origin domain and then of the rest of the chromosome. Low-copy number plasmids rely on specialized partitioning systems, which in some cases use a mechanism that show striking similarity to eukaryotic DNA segregation. Overall, there have been multiple systems implicated in the dynamic transport of DNA cargo to a new cellular position during the cell cycle but most seem to share a common initial DNA partitioning step, involving the formation of a nucleoprotein complex called the segrosome. The particular features and complex topologies of individual segrosomes depend on both the nature of the DNA binding protein involved and on the recognized centromeric DNA sequence, both of which vary across systems. The combination of in vivo and in vitro approaches, with structural biology has significantly furthered our understanding of the mechanisms underlying DNA trafficking in bacteria. Here, I discuss recent advances and the molecular details of the DNA segregation machinery, focusing on the formation of the segrosome complex.
Keywords: DNA segregation; ParB; ParR; TubR; nucleoprotein complex; partitioning complex; partitioning systems; segrosome.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources