

Tcf7l2/Tcf4 Transcriptional Repressor Function Requires HDAC Activity in the Developing Vertebrate CNS
- PMID: 27668865
- PMCID: PMC5036887
- DOI: 10.1371/journal.pone.0163267
Tcf7l2/Tcf4 Transcriptional Repressor Function Requires HDAC Activity in the Developing Vertebrate CNS
Abstract
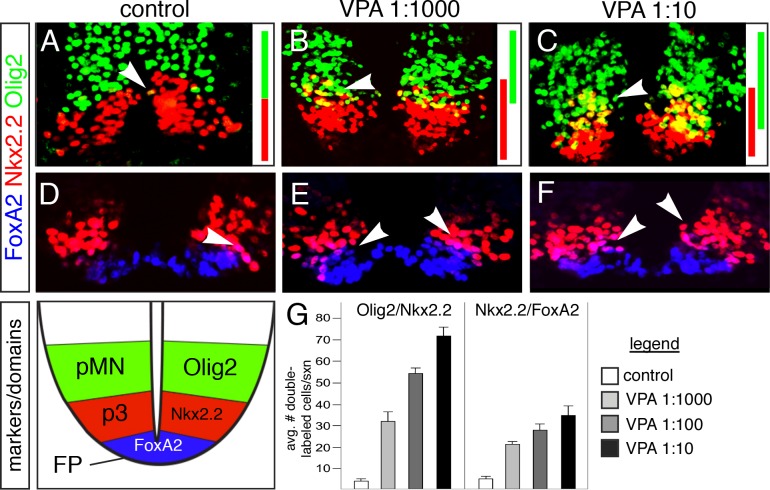
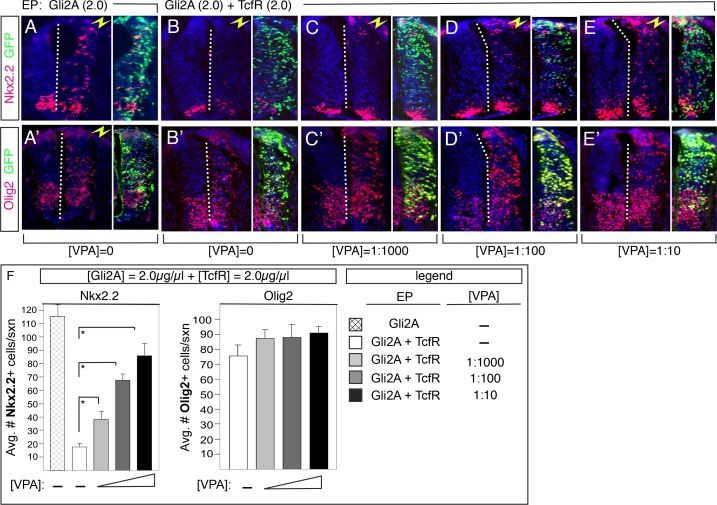
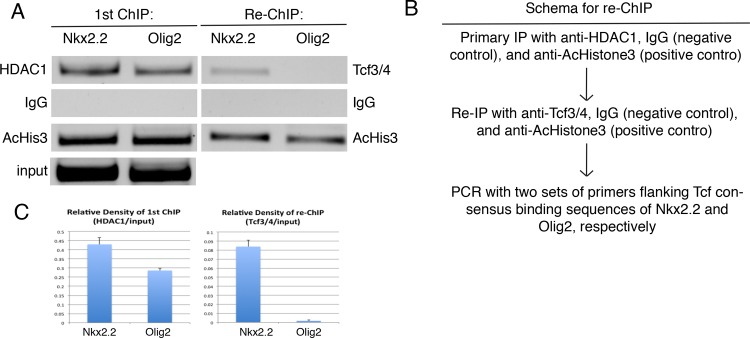
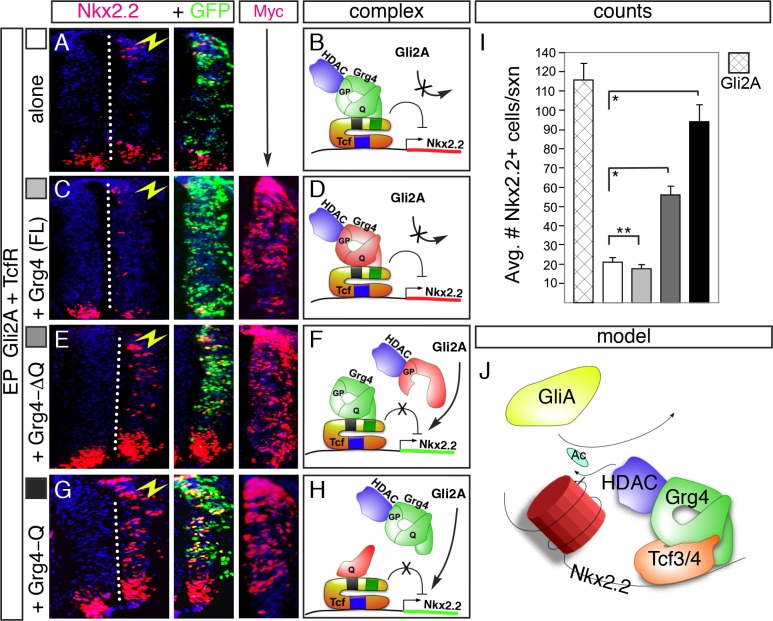
The generation of functionally distinct neuronal subtypes within the vertebrate central nervous system (CNS) requires the precise regulation of progenitor gene expression in specific neuronal territories during early embryogenesis. Accumulating evidence has implicated histone deacetylase (HDAC) proteins in cell specification, proliferation, and differentiation in diverse embryonic and adult tissues. However, although HDAC proteins have shown to be expressed in the developing vertebrate neural tube, their specific role in CNS neural progenitor fate specification remains unclear. Prior work from our lab showed that the Tcf7l2/Tcf4 transcription factor plays a key role in ventral progenitor lineage segregation by differential repression of two key specification factors, Nkx2.2 and Olig2. In this study, we found that administration of HDAC inhibitors (Valproic Acid (VPA), Trichostatin-A (TSA), or sodium butyrate) in chick embryos in ovo disrupted normal progenitor gene segregation in the developing neural tube, indicating that HDAC activity is required for this process. Further, using functional and pharmacological approaches in vivo, we found that HDAC activity is required for the differential repression of Nkx2.2 and Olig2 by Tcf7l2/Tcf4. Finally, using dominant-negative functional assays, we provide evidence that Tcf7l2/Tcf4 repression also requires Gro/TLE/Grg co-repressor factors. Together, our data support a model where the transcriptional repressor activity of Tcf7l2/Tcf4 involves functional interactions with both HDAC and Gro/TLE/Grg co-factors at specific target gene regulatory elements in the developing neural tube, and that this activity is required for the proper segregation of the Nkx2.2 (p3) and Olig2 (pMN) expressing cells from a common progenitor pool.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Histone deacetylase inhibitor Trichostatin A induces neural tube defects and promotes neural crest specification in the chicken neural tube.Differentiation. 2013 Jan;85(1-2):55-66. doi: 10.1016/j.diff.2012.12.001. Epub 2013 Jan 16. Differentiation. 2013. PMID: 23328540
-
The neuron-restrictive silencer element-neuron-restrictive silencer factor system regulates basal and endothelin 1-inducible atrial natriuretic peptide gene expression in ventricular myocytes.Mol Cell Biol. 2001 Mar;21(6):2085-97. doi: 10.1128/MCB.21.6.2085-2097.2001. Mol Cell Biol. 2001. PMID: 11238943 Free PMC article.
-
Stra13 expression is associated with growth arrest and represses transcription through histone deacetylase (HDAC)-dependent and HDAC-independent mechanisms.Proc Natl Acad Sci U S A. 2000 Apr 11;97(8):4058-63. doi: 10.1073/pnas.070526297. Proc Natl Acad Sci U S A. 2000. PMID: 10737769 Free PMC article.
-
p21Waf1/Cip1 is a common target induced by short-chain fatty acid HDAC inhibitors (valproic acid, tributyrin and sodium butyrate) in neuroblastoma cells.Oncol Rep. 2005 Jun;13(6):1139-44. Oncol Rep. 2005. PMID: 15870934
-
Inhibition of histone deacetylase activity on specific embryonic tissues as a new mechanism for teratogenicity.Birth Defects Res B Dev Reprod Toxicol. 2005 Oct;74(5):392-8. doi: 10.1002/bdrb.20053. Birth Defects Res B Dev Reprod Toxicol. 2005. PMID: 16193500
Cited by
-
Whole Exome Sequencing Identifies APCDD1 and HDAC5 Genes as Potentially Cancer Predisposing in Familial Colorectal Cancer.Int J Mol Sci. 2021 Feb 12;22(4):1837. doi: 10.3390/ijms22041837. Int J Mol Sci. 2021. PMID: 33673279 Free PMC article. Clinical Trial.
-
Opposing roles of TCF7/LEF1 and TCF7L2 in cyclin D2 and Bmp4 expression and cardiomyocyte cell cycle control during late heart development.Lab Invest. 2019 Jun;99(6):807-818. doi: 10.1038/s41374-019-0204-2. Epub 2019 Feb 18. Lab Invest. 2019. PMID: 30778164 Free PMC article.
-
Characterization of Coding/Noncoding Variants for SHROOM3 in Patients with CKD.J Am Soc Nephrol. 2018 May;29(5):1525-1535. doi: 10.1681/ASN.2017080856. Epub 2018 Feb 23. J Am Soc Nephrol. 2018. PMID: 29476007 Free PMC article.
-
Abundant repressor binding sites in human enhancers are associated with the fine-tuning of gene regulation.iScience. 2024 Dec 20;28(1):111658. doi: 10.1016/j.isci.2024.111658. eCollection 2025 Jan 17. iScience. 2024. PMID: 39868043 Free PMC article.
-
Histone Deacetylase Inhibitors Impair Glioblastoma Cell Motility and Proliferation.Cancers (Basel). 2022 Apr 9;14(8):1897. doi: 10.3390/cancers14081897. Cancers (Basel). 2022. PMID: 35454804 Free PMC article.
References
-
- Dessaud E, Yang LL, Hill K, Cox B, Ulloa F, et al. (2007) Interpretation of the sonic hedgehog morphogen gradient by a temporal adaptation mechanism. Nature 450: 717–720. - PubMed
-
- Rowitch DH (2004) Glial specification in the vertebrate neural tube. Nat Rev Neurosci 5: 409–419. - PubMed
-
- Briscoe J, Pierani A, Jessell TM, Ericson J (2000) A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube. Cell 101: 435–445. - PubMed
-
- Muhr J, Andersson E, Persson M, Jessell TM, Ericson J (2001) Groucho-mediated transcriptional repression establishes progenitor cell pattern and neuronal fate in the ventral neural tube. Cell 104: 861–873. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
