

A visual circuit uses complementary mechanisms to support transient and sustained pupil constriction
- PMID: 27669145
- PMCID: PMC5079752
- DOI: 10.7554/eLife.15392
A visual circuit uses complementary mechanisms to support transient and sustained pupil constriction
Abstract
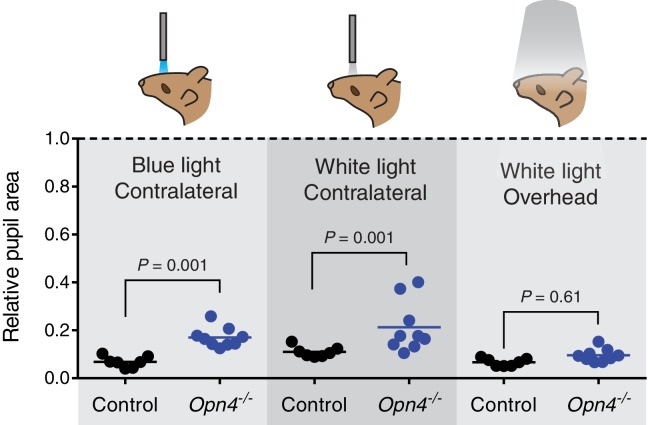
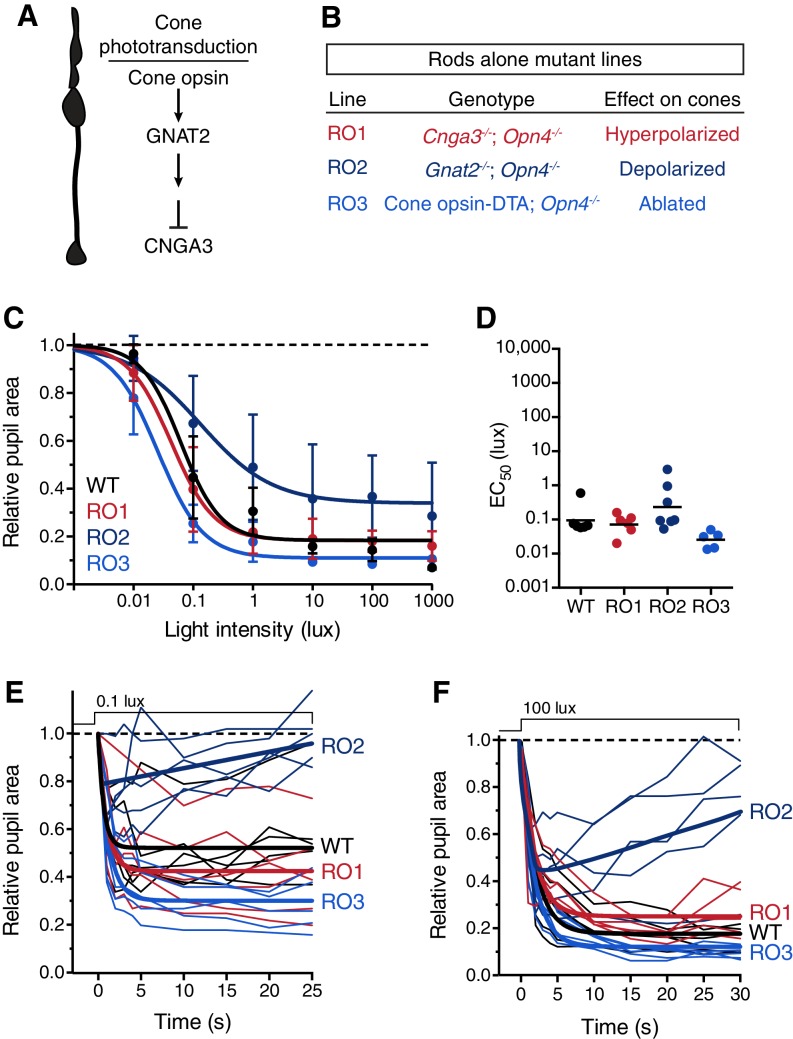
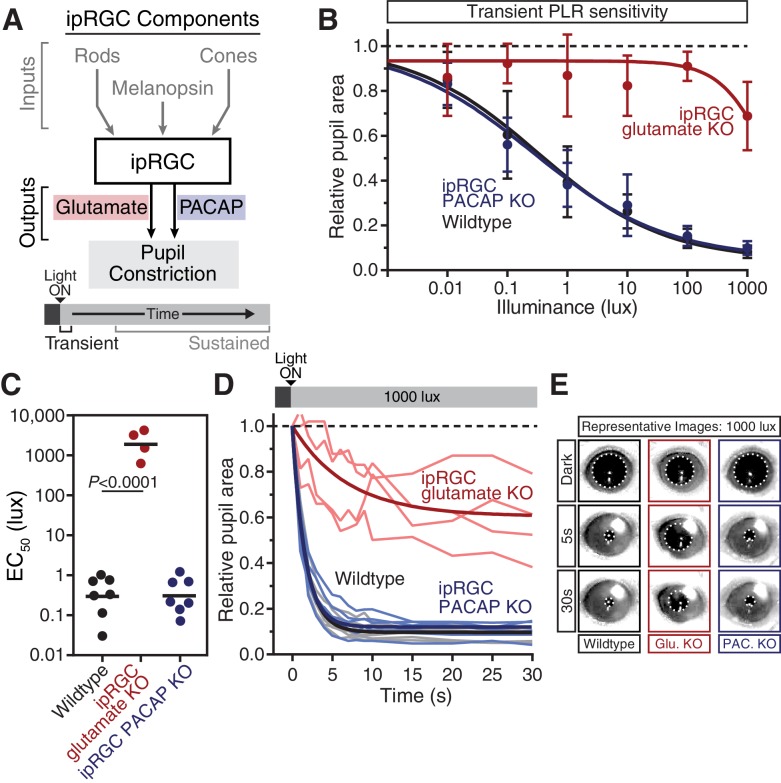
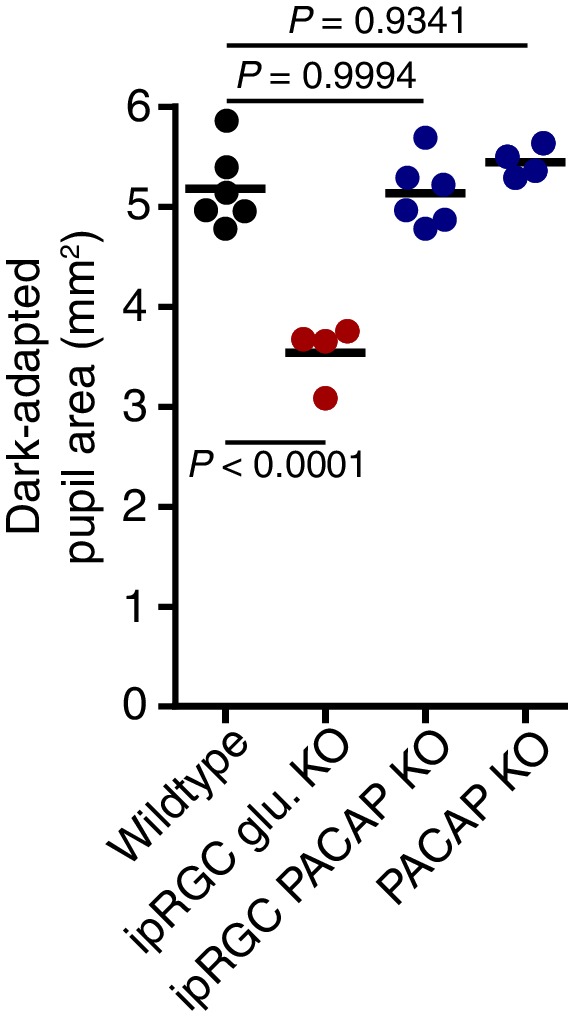
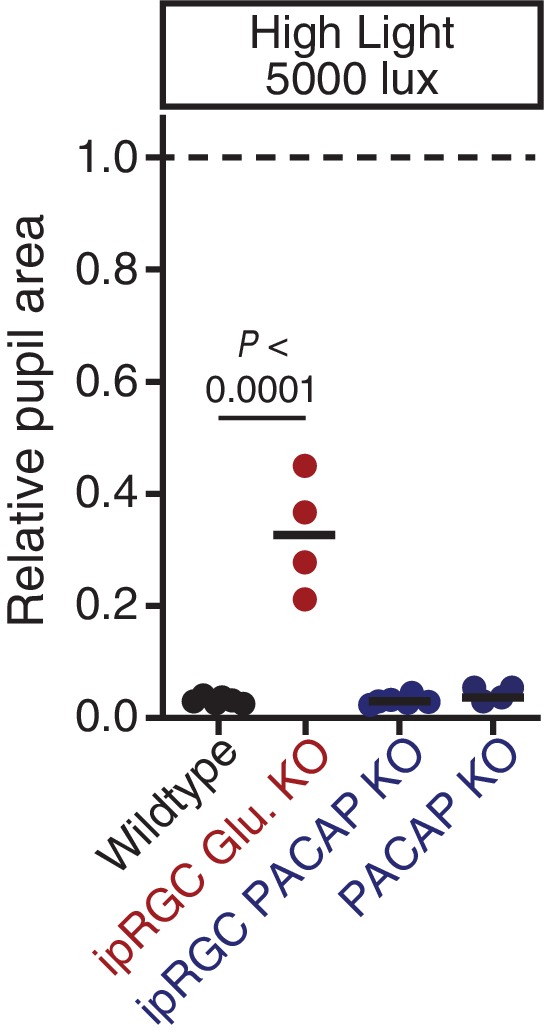
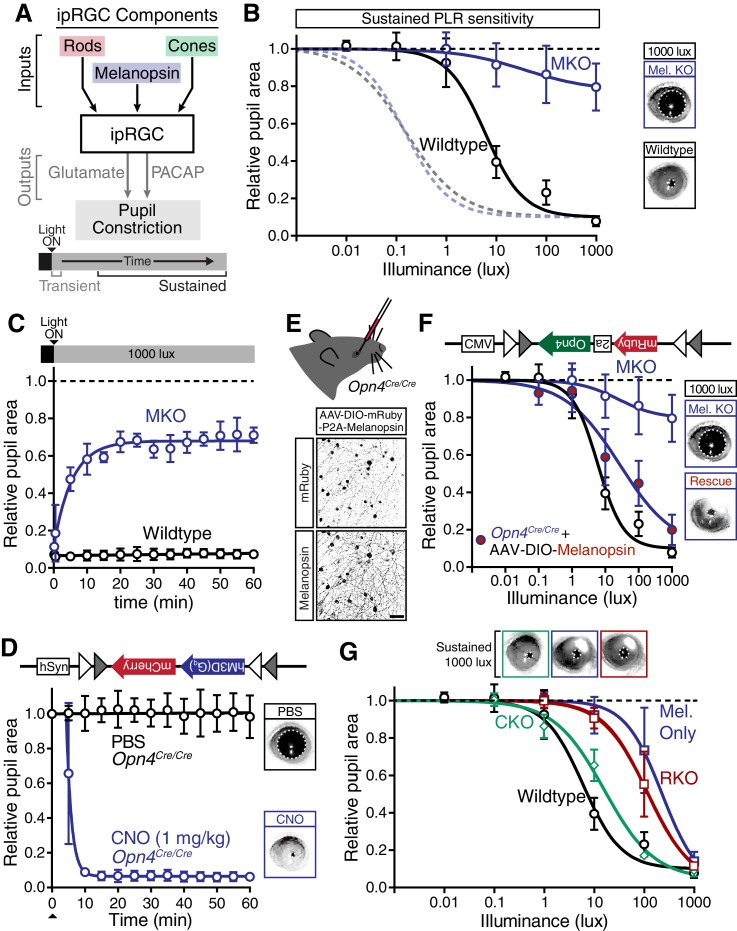
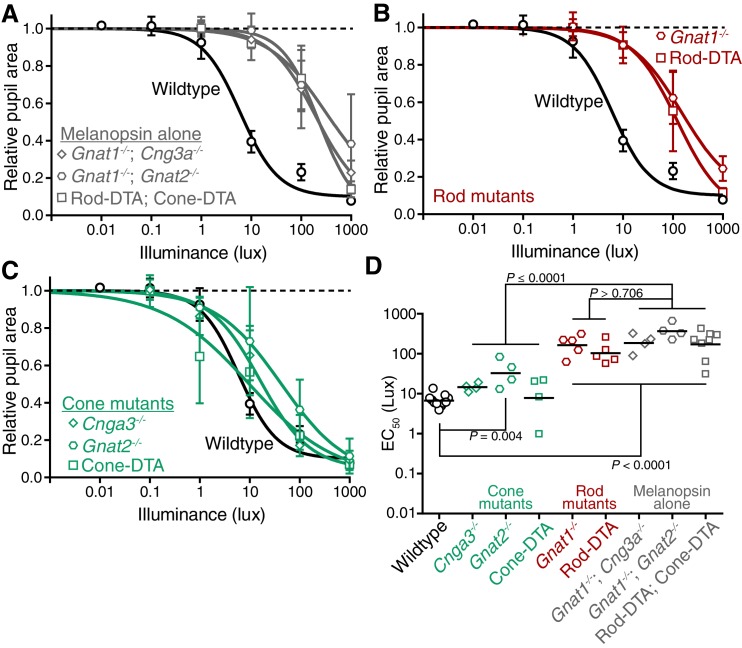
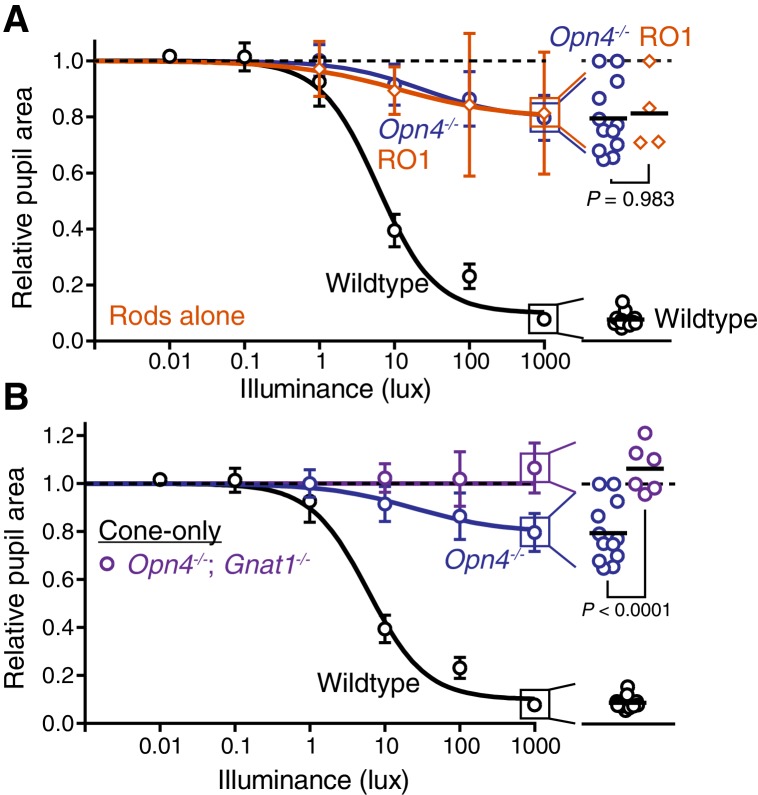
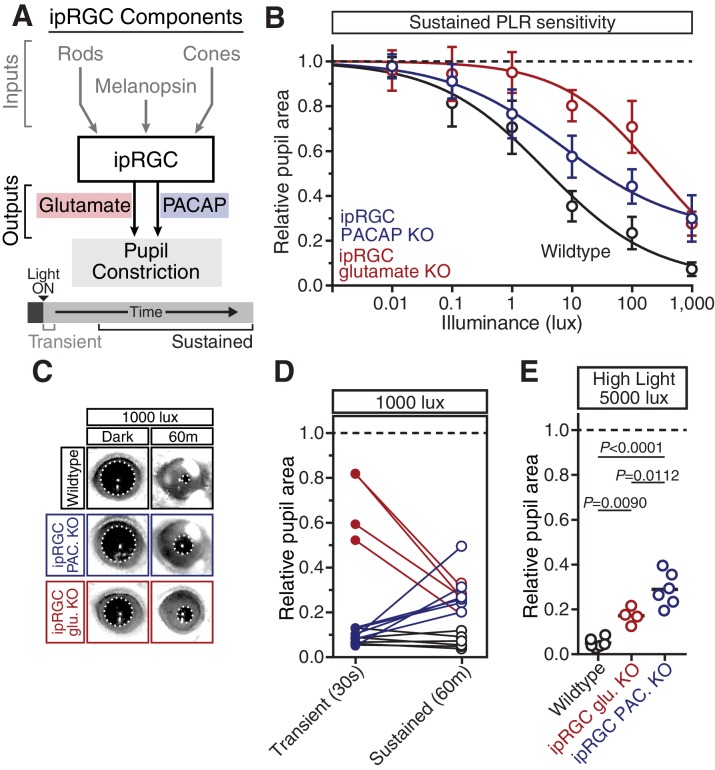
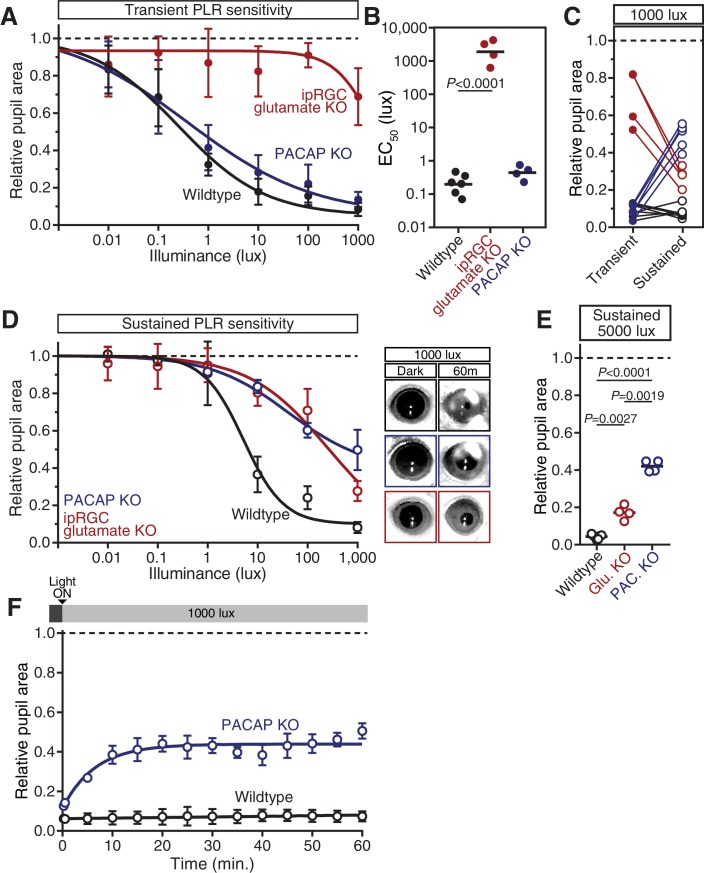
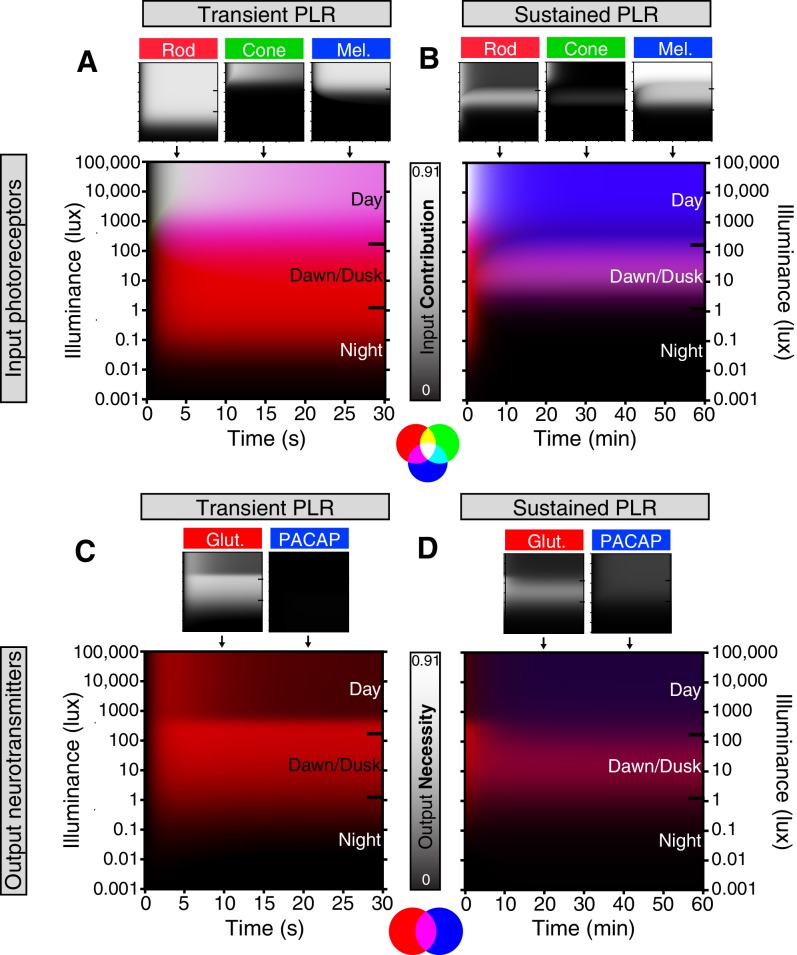
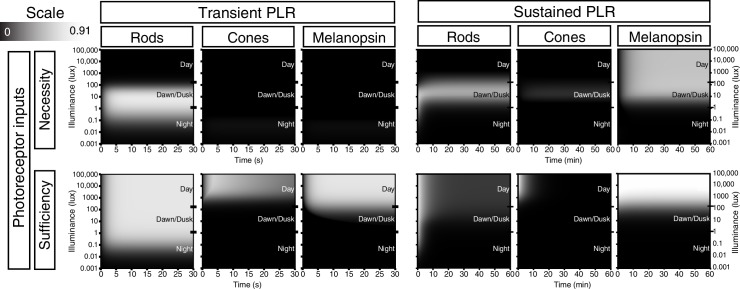
Rapid and stable control of pupil size in response to light is critical for vision, but the neural coding mechanisms remain unclear. Here, we investigated the neural basis of pupil control by monitoring pupil size across time while manipulating each photoreceptor input or neurotransmitter output of intrinsically photosensitive retinal ganglion cells (ipRGCs), a critical relay in the control of pupil size. We show that transient and sustained pupil responses are mediated by distinct photoreceptors and neurotransmitters. Transient responses utilize input from rod photoreceptors and output by the classical neurotransmitter glutamate, but adapt within minutes. In contrast, sustained responses are dominated by non-conventional signaling mechanisms: melanopsin phototransduction in ipRGCs and output by the neuropeptide PACAP, which provide stable pupil maintenance across the day. These results highlight a temporal switch in the coding mechanisms of a neural circuit to support proper behavioral dynamics.
Keywords: ipRGC; melanopsin; mouse; neuropeptides; neuroscience; neurotransmitters; retinal circuitry; vision.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
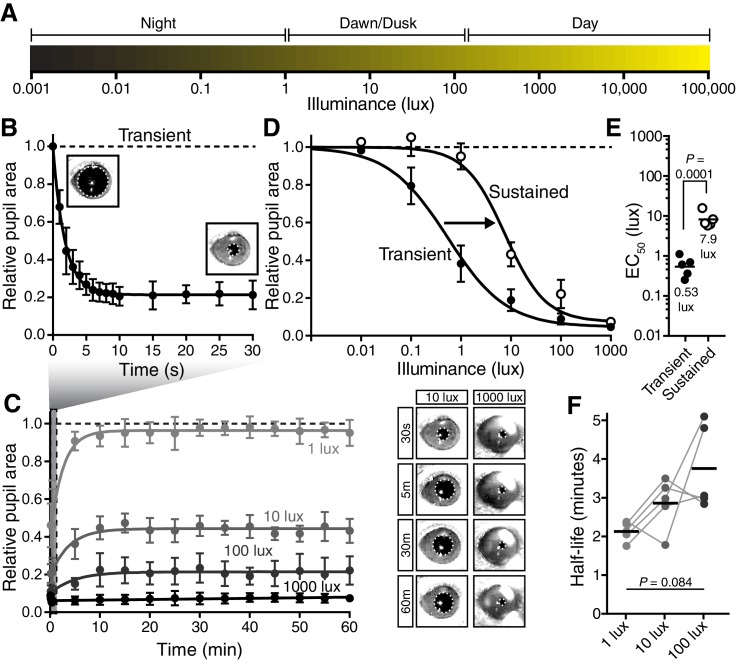
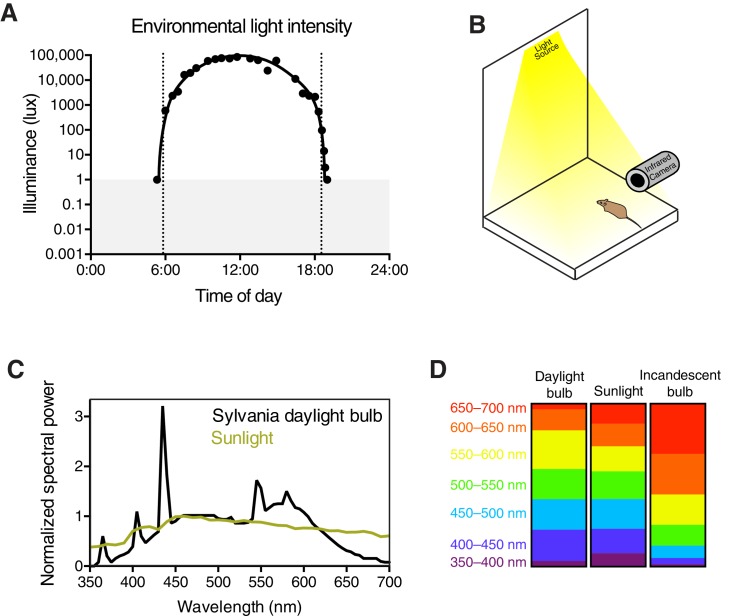
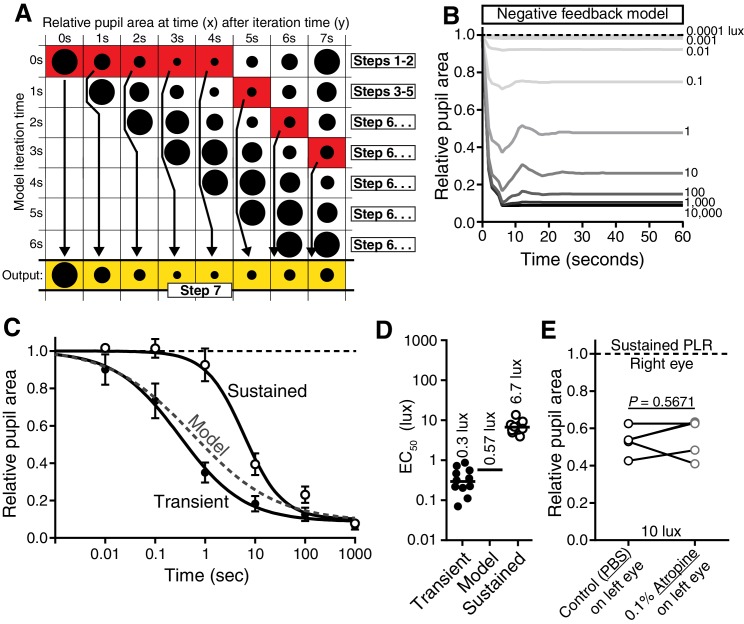

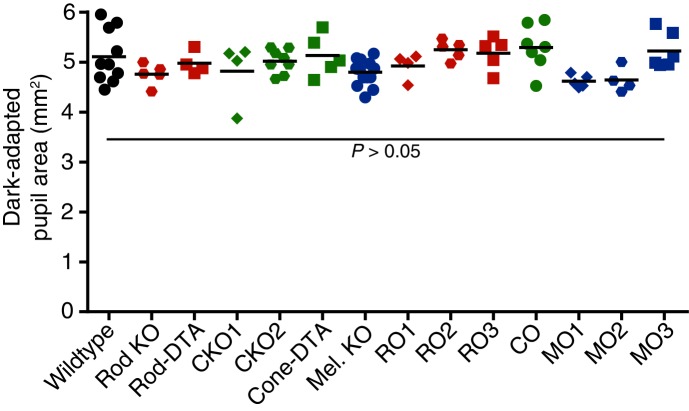





Similar articles
-
Temporal characteristics of melanopsin inputs to the human pupil light reflex.Vision Res. 2015 Feb;107:58-66. doi: 10.1016/j.visres.2014.12.001. Epub 2014 Dec 10. Vision Res. 2015. PMID: 25497360 Free PMC article.
-
A five-primary photostimulator suitable for studying intrinsically photosensitive retinal ganglion cell functions in humans.J Vis. 2015 Jan 26;15(1):15.1.27. doi: 10.1167/15.1.27. J Vis. 2015. PMID: 25624466 Free PMC article.
-
Intrinsically photosensitive retinal ganglion cells.J Neuroophthalmol. 2007 Sep;27(3):195-204. doi: 10.1097/WNO.0b013e31814b1df9. J Neuroophthalmol. 2007. PMID: 17895821 Review.
-
Melanopsin and Cone Photoreceptor Inputs to the Afferent Pupil Light Response.Front Neurol. 2019 May 22;10:529. doi: 10.3389/fneur.2019.00529. eCollection 2019. Front Neurol. 2019. PMID: 31191431 Free PMC article.
-
[Pupil and melanopsin photoreception].Nippon Ganka Gakkai Zasshi. 2013 Mar;117(3):246-68; discussion 269. Nippon Ganka Gakkai Zasshi. 2013. PMID: 23631256 Review. Japanese.
Cited by
-
Diversity of intrinsically photosensitive retinal ganglion cells: circuits and functions.Cell Mol Life Sci. 2021 Feb;78(3):889-907. doi: 10.1007/s00018-020-03641-5. Epub 2020 Sep 23. Cell Mol Life Sci. 2021. PMID: 32965515 Free PMC article. Review.
-
Sustained effects of prior red light on pupil diameter and vigilance during subsequent darkness.Proc Biol Sci. 2018 Jul 18;285(1883):20180989. doi: 10.1098/rspb.2018.0989. Proc Biol Sci. 2018. PMID: 30051840 Free PMC article.
-
Dynamics of Non-visual Responses in Humans: As Fast as Lightning?Front Neurosci. 2019 Mar 5;13:126. doi: 10.3389/fnins.2019.00126. eCollection 2019. Front Neurosci. 2019. PMID: 30890907 Free PMC article.
-
Melanopsin-mediated optical entrainment regulates circadian rhythms in vertebrates.Commun Biol. 2023 Oct 18;6(1):1054. doi: 10.1038/s42003-023-05432-7. Commun Biol. 2023. PMID: 37853054 Free PMC article. Review.
-
Retinal pigment epithelium 65 kDa protein (RPE65): An update.Prog Retin Eye Res. 2022 May;88:101013. doi: 10.1016/j.preteyeres.2021.101013. Epub 2021 Oct 2. Prog Retin Eye Res. 2022. PMID: 34607013 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
