

Value, search, persistence and model updating in anterior cingulate cortex
- PMID: 27669988
- PMCID: PMC7116891
- DOI: 10.1038/nn.4382
Value, search, persistence and model updating in anterior cingulate cortex
Abstract
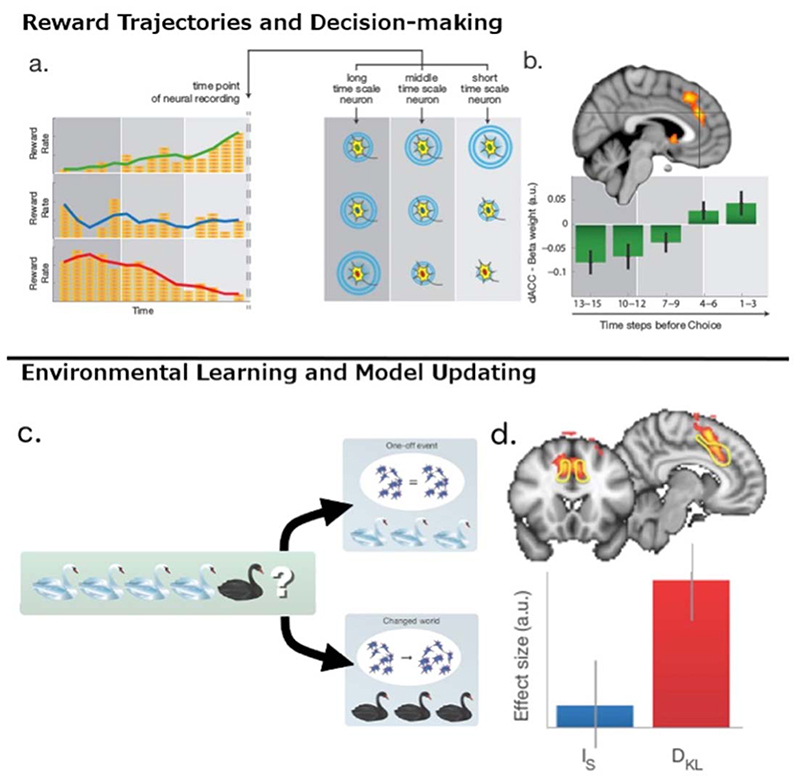
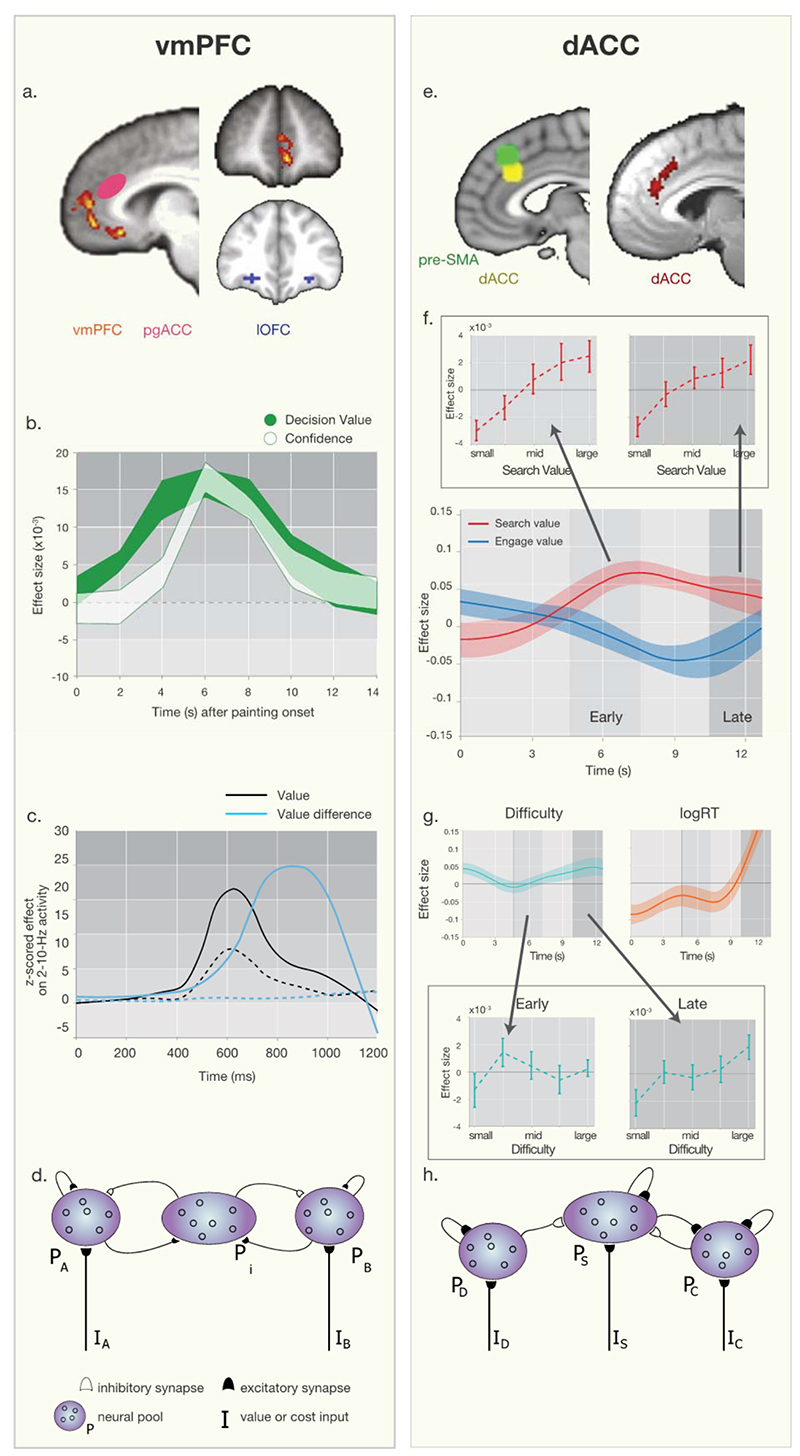
Dorsal anterior cingulate cortex (dACC) carries a wealth of value-related information necessary for regulating behavioral flexibility and persistence. It signals error and reward events informing decisions about switching or staying with current behavior. During decision-making, it encodes the average value of exploring alternative choices (search value), even after controlling for response selection difficulty, and during learning, it encodes the degree to which internal models of the environment and current task must be updated. dACC value signals are derived in part from the history of recent reward integrated simultaneously over multiple time scales, thereby enabling comparison of experience over the recent and extended past. Such ACC signals may instigate attentionally demanding and difficult processes such as behavioral change via interactions with prefrontal cortex. However, the signal in dACC that instigates behavioral change need not itself be a conflict or difficulty signal.
Figures
Comment in
-
Dorsal anterior cingulate: a Rorschach test for cognitive neuroscience.Nat Neurosci. 2016 Sep 27;19(10):1278-9. doi: 10.1038/nn.4387. Nat Neurosci. 2016. PMID: 27669987 No abstract available.
References
-
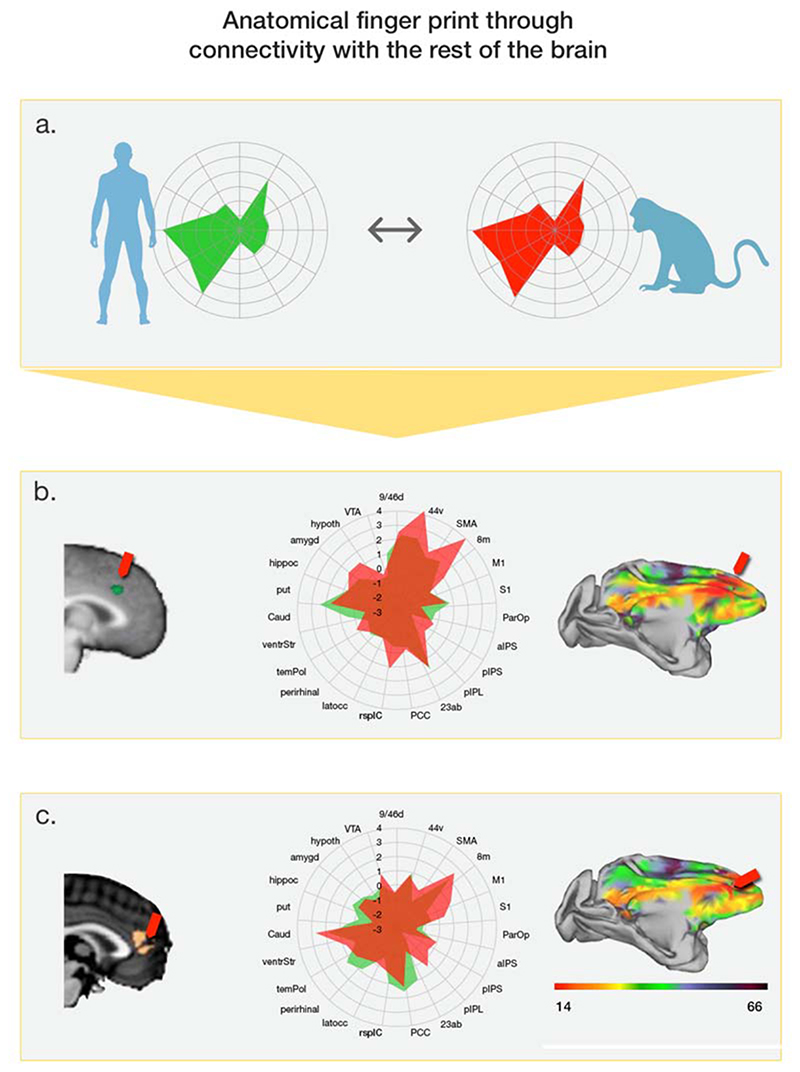
- Neubert FX, Mars RB, Sallet J, Rushworth MF. Connectivity reveals relationship of brain areas for reward-guided learning and decision making in human and monkey frontal cortex. Proceedings of the National Academy of Sciences of the United States of America. 2015 doi: 10.1073/pnas.1410767112. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
