

LTD-like molecular pathways in developmental synaptic pruning
- PMID: 27669991
- PMCID: PMC5070480
- DOI: 10.1038/nn.4389
LTD-like molecular pathways in developmental synaptic pruning
Abstract
In long-term depression (LTD) at synapses in the adult brain, synaptic strength is reduced in an experience-dependent manner. LTD thus provides a cellular mechanism for information storage in some forms of learning. A similar activity-dependent reduction in synaptic strength also occurs in the developing brain and there provides an essential step in synaptic pruning and the postnatal development of neural circuits. Here we review evidence suggesting that LTD and synaptic pruning share components of their underlying molecular machinery and may thus represent two developmental stages of the same type of synaptic modulation that serve different, but related, functions in neural circuit plasticity. We also assess the relationship between LTD and synaptic pruning in the context of recent findings of LTD dysregulation in several mouse models of autism spectrum disorder (ASD) and discuss whether LTD deficits can indicate impaired pruning processes that are required for proper brain development.
Figures
Similar articles
-
Deregulation of synaptic plasticity in autism.Neurosci Lett. 2019 Jan 1;688:58-61. doi: 10.1016/j.neulet.2018.02.003. Epub 2018 Feb 5. Neurosci Lett. 2019. PMID: 29421544 Review.
-
Microglia in the pathogenesis of autism spectrum disorders.Neurosci Res. 2015 Nov;100:1-5. doi: 10.1016/j.neures.2015.06.005. Epub 2015 Jun 25. Neurosci Res. 2015. PMID: 26116891 Review.
-
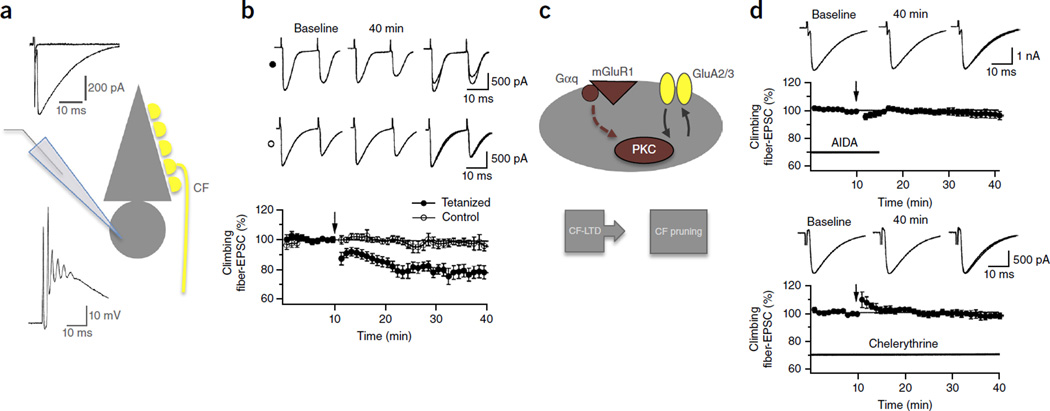
Long-Term Depression of Intrinsic Excitability Accompanied by Synaptic Depression in Cerebellar Purkinje Cells.J Neurosci. 2017 Jun 7;37(23):5659-5669. doi: 10.1523/JNEUROSCI.3464-16.2017. Epub 2017 May 11. J Neurosci. 2017. PMID: 28495974 Free PMC article.
-
The neurobiological bases of autism spectrum disorders: the R451C-neuroligin 3 mutation hampers the expression of long-term synaptic depression in the dorsal striatum.Eur J Neurosci. 2018 Mar;47(6):701-708. doi: 10.1111/ejn.13705. Epub 2017 Oct 4. Eur J Neurosci. 2018. PMID: 28921757
-
Long-term depression and other synaptic plasticity in the cerebellum.Proc Jpn Acad Ser B Phys Biol Sci. 2013;89(5):183-95. doi: 10.2183/pjab.89.183. Proc Jpn Acad Ser B Phys Biol Sci. 2013. PMID: 23666089 Free PMC article. Review.
Cited by
-
c-Abl Deficiency Provides Synaptic Resiliency Against Aβ-Oligomers.Front Cell Neurosci. 2019 Nov 26;13:526. doi: 10.3389/fncel.2019.00526. eCollection 2019. Front Cell Neurosci. 2019. PMID: 31849613 Free PMC article.
-
Aberrant Synaptic PTEN in Symptomatic Alzheimer's Patients May Link Synaptic Depression to Network Failure.Front Synaptic Neurosci. 2021 May 11;13:683290. doi: 10.3389/fnsyn.2021.683290. eCollection 2021. Front Synaptic Neurosci. 2021. PMID: 34045952 Free PMC article.
-
Removing 4E-BP Enables Synapses to Refine without Postsynaptic Activity.Cell Rep. 2018 Apr 3;23(1):11-22. doi: 10.1016/j.celrep.2018.03.040. Cell Rep. 2018. PMID: 29617653 Free PMC article.
-
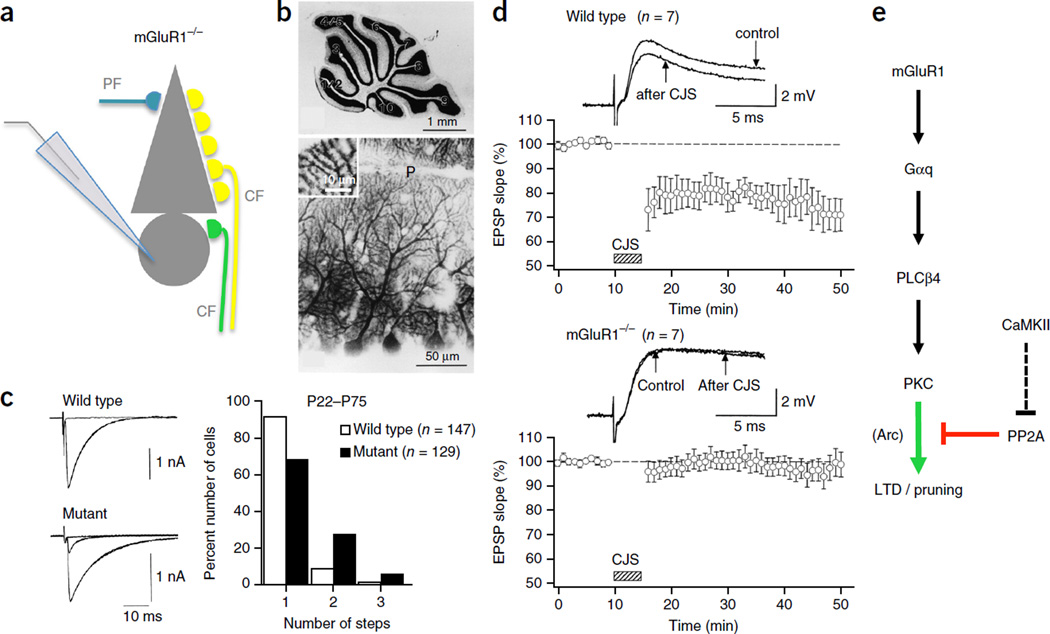
Type-1 metabotropic glutamate receptor signaling in cerebellar Purkinje cells in health and disease.F1000Res. 2017 Apr 4;6:416. doi: 10.12688/f1000research.10485.1. eCollection 2017. F1000Res. 2017. PMID: 28435670 Free PMC article. Review.
-
The impact of mild episodic ketosis on microglia and hippocampal long-term depression in 5xFAD mice.FASEB Bioadv. 2024 Oct 23;6(12):581-596. doi: 10.1096/fba.2024-00123. eCollection 2024 Dec. FASEB Bioadv. 2024. PMID: 39650227 Free PMC article.
References
-
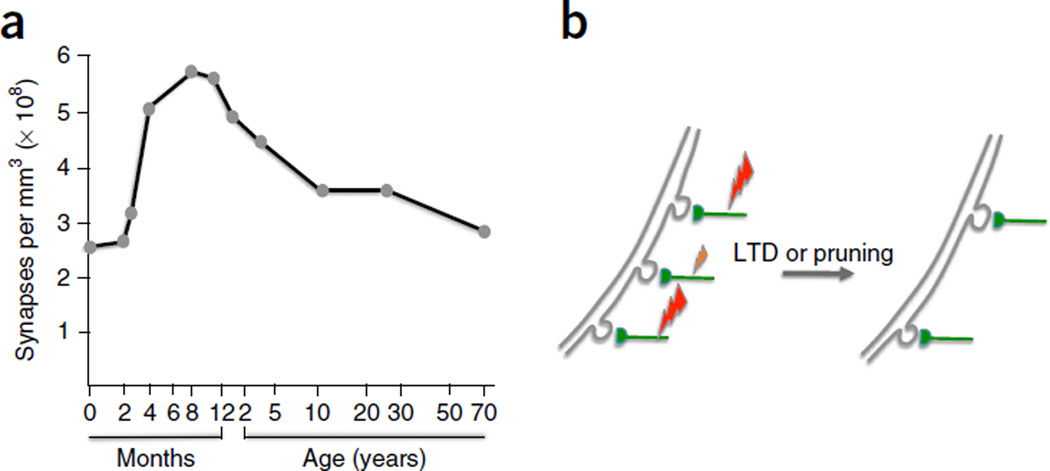
- Huttenlocher PR. Morphometric study of human cerebral cortex development. Neuropsychologia. 1990;28:517–527. - PubMed
-
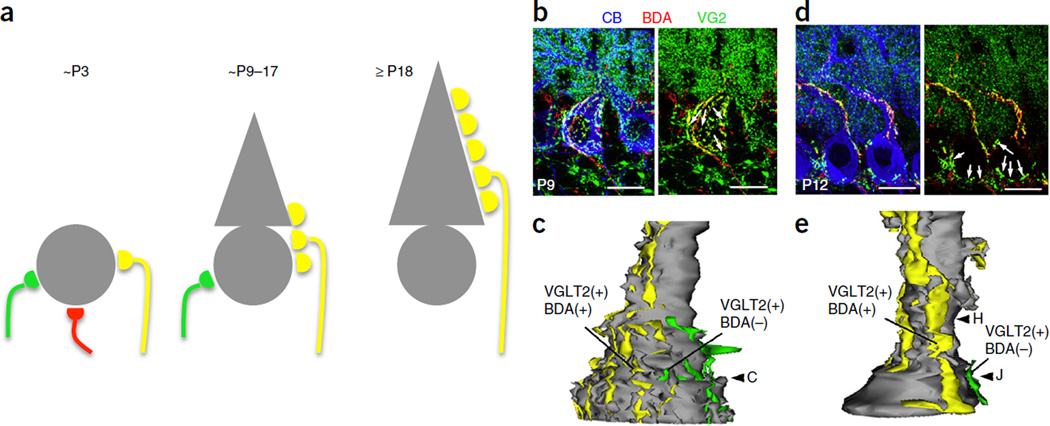
- Tapia JC, et al. Pervasive synaptic branch removal in the mammalian neuromuscular system at birth. Neuron. 2012;74:816–829. - PubMed
-
- Colman H, Nabekura J, Lichtman JW. Alterations in synaptic strength preceding axon withdrawal. Science. 1997;275:356–361. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
