

Flow-dependent YAP/TAZ activities regulate endothelial phenotypes and atherosclerosis
- PMID: 27671657
- PMCID: PMC5068257
- DOI: 10.1073/pnas.1613121113
Flow-dependent YAP/TAZ activities regulate endothelial phenotypes and atherosclerosis
Abstract
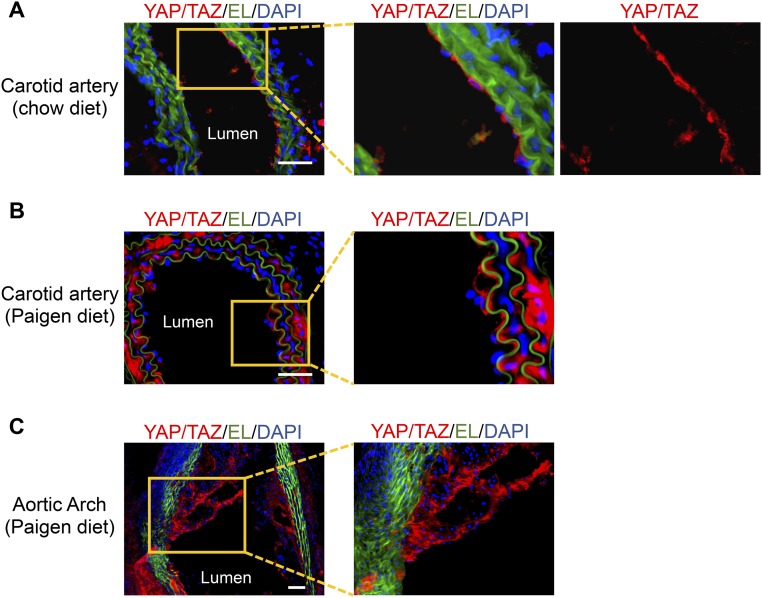
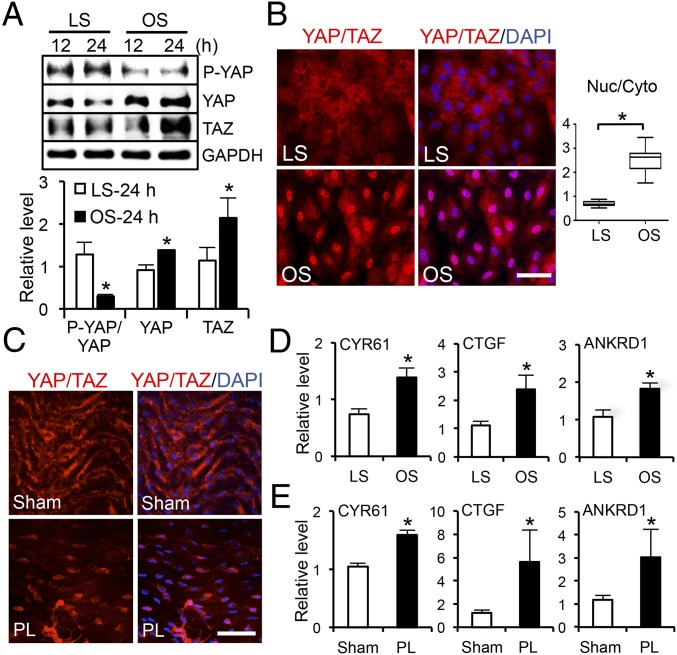
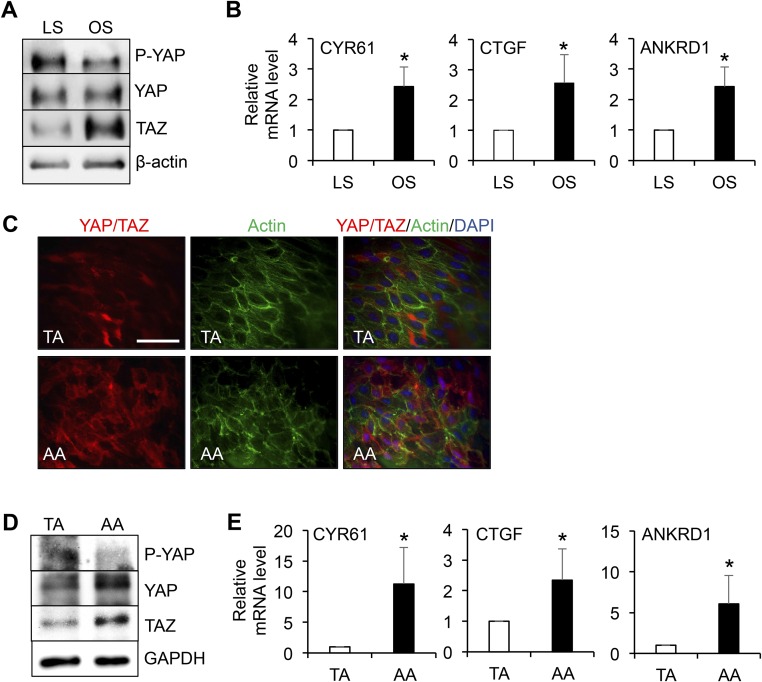
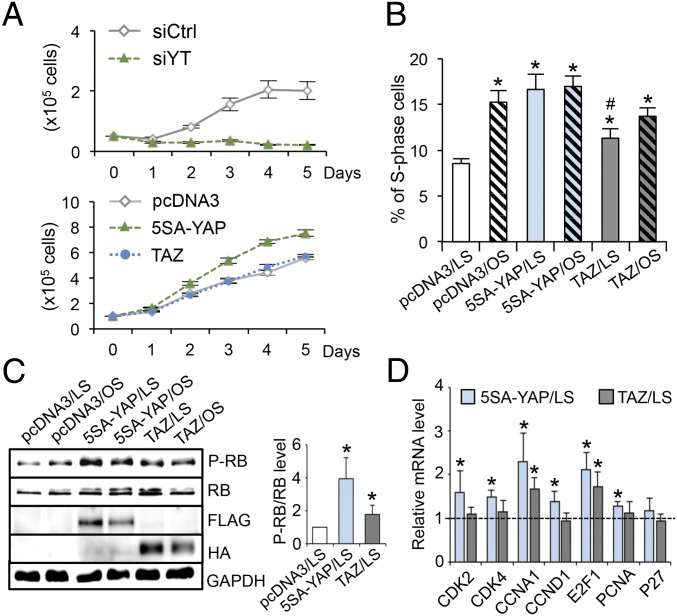
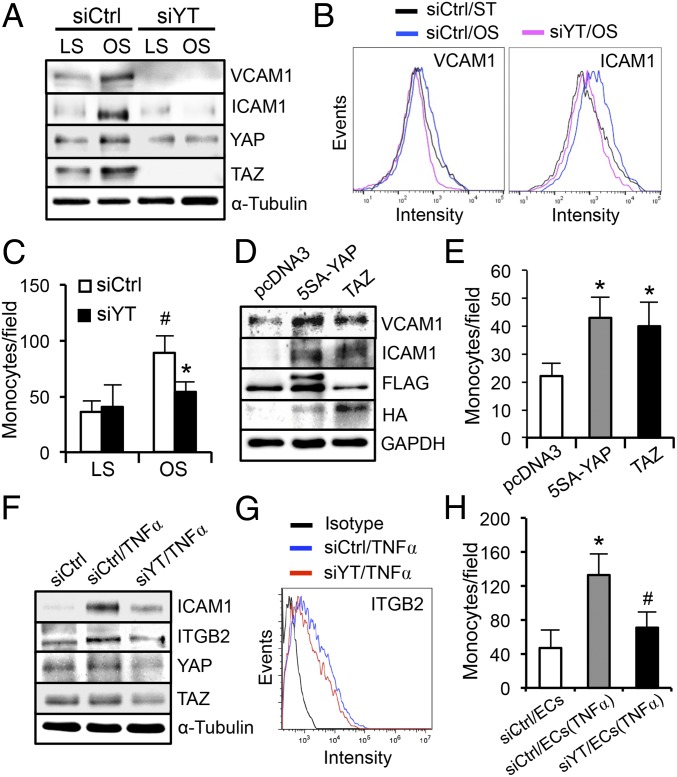
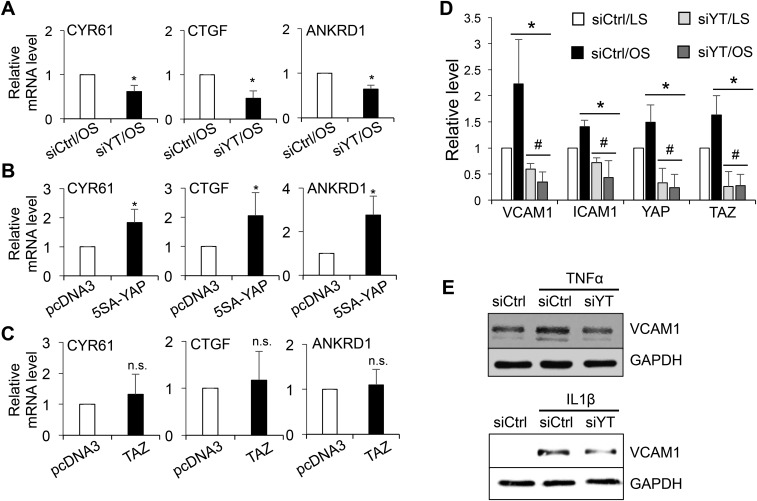
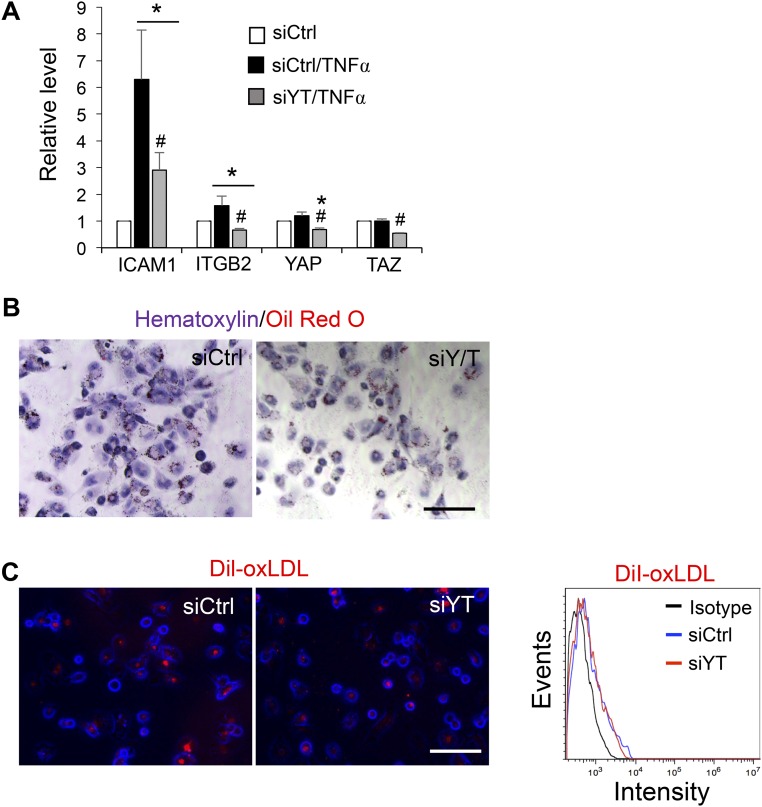
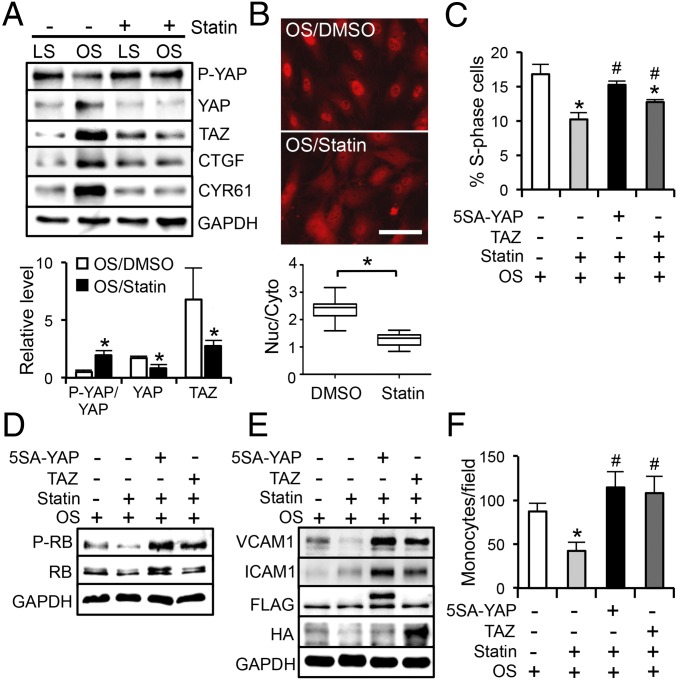
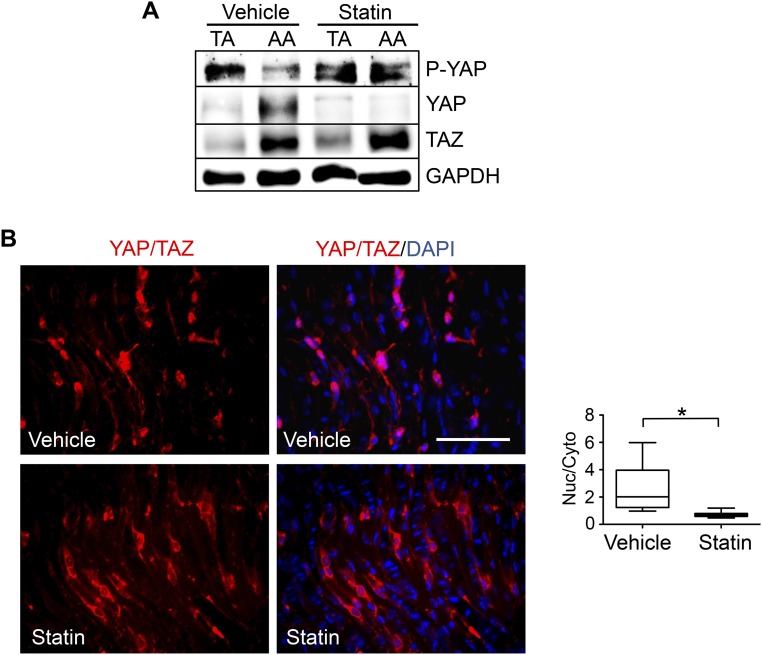
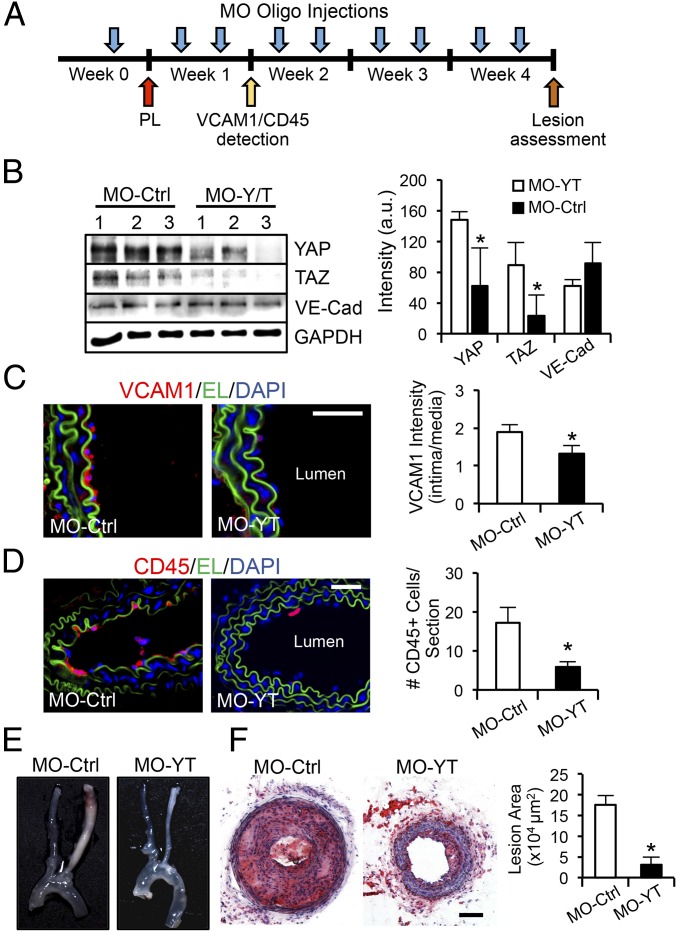
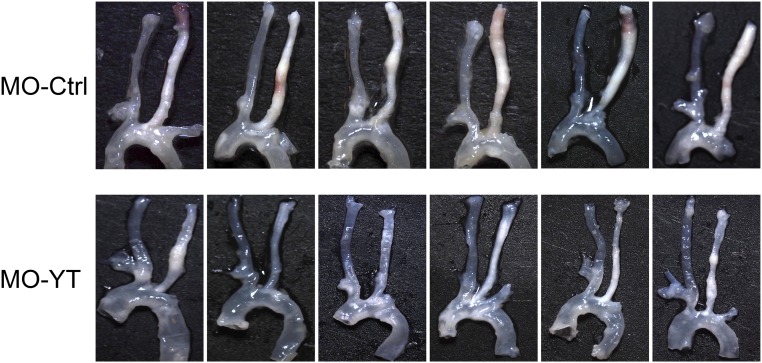
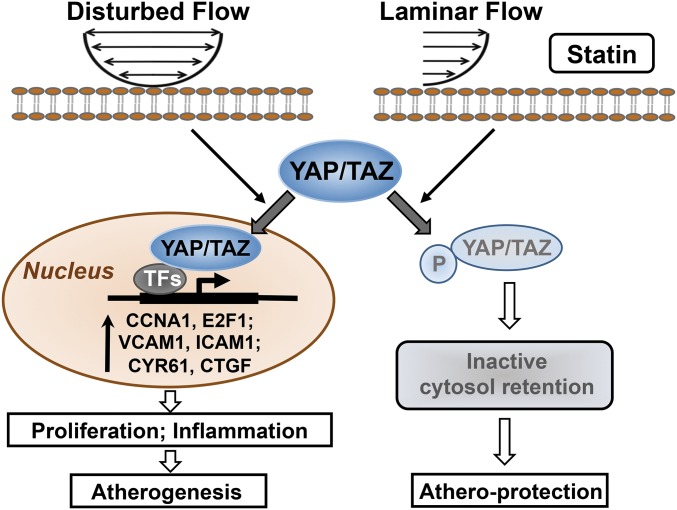
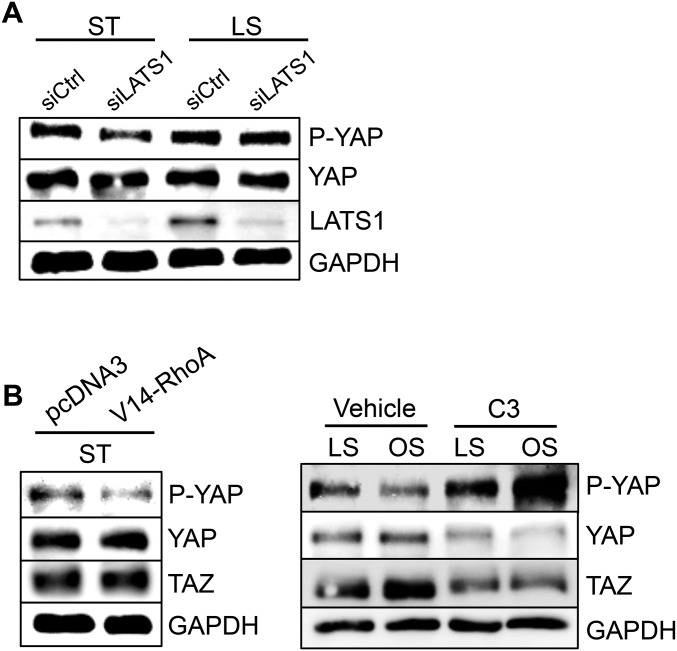
The focal nature of atherosclerotic lesions suggests an important role of local hemodynamic environment. Recent studies have demonstrated significant roles of Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ) in mediating mechanotransduction and vascular homeostasis. The objective of this study is to investigate the functional role of YAP/TAZ in the flow regulation of atheroprone endothelial phenotypes and the consequential development of atherosclerotic lesions. We found that exposure of cultured endothelial cells (ECs) to the atheroprone disturbed flow resulted in YAP/TAZ activation and translocation into EC nucleus to up-regulate the target genes, including cysteine-rich angiogenic inducer 61 (CYR61), connective tissue growth factor (CTGF), and ankyrin repeat domain 1 (ANKRD1). In contrast, the athero-protective laminar flow suppressed YAP/TAZ activities. En face analysis of mouse arteries demonstrated an increased nuclear localization of YAP/TAZ and elevated levels of the target genes in the endothelium in atheroprone areas compared with athero-protective areas. YAP/TAZ knockdown significantly attenuated the disturbed flow induction of EC proliferative and proinflammatory phenotypes, whereas overexpression of constitutively active YAP was sufficient to promote EC proliferation and inflammation. In addition, treatment with statin, an antiatherosclerotic drug, inhibited YAP/TAZ activities to diminish the disturbed flow-induced proliferation and inflammation. In vivo blockade of YAP/TAZ translation by morpholino oligos significantly reduced endothelial inflammation and the size of atherosclerotic lesions. Our results demonstrate a critical role of the activation of YAP/TAZ by disturbed flow in promoting atheroprone phenotypes and atherosclerotic lesion development. Therefore, inhibition of YAP/TAZ activation is a promising athero-protective therapeutic strategy.
Keywords: atherogenesis; disturbed flow; endothelial cells; mechanotransduction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
