

Validation of CoaBC as a Bactericidal Target in the Coenzyme A Pathway of Mycobacterium tuberculosis
- PMID: 27676316
- PMCID: PMC5153693
- DOI: 10.1021/acsinfecdis.6b00150
Validation of CoaBC as a Bactericidal Target in the Coenzyme A Pathway of Mycobacterium tuberculosis
Abstract
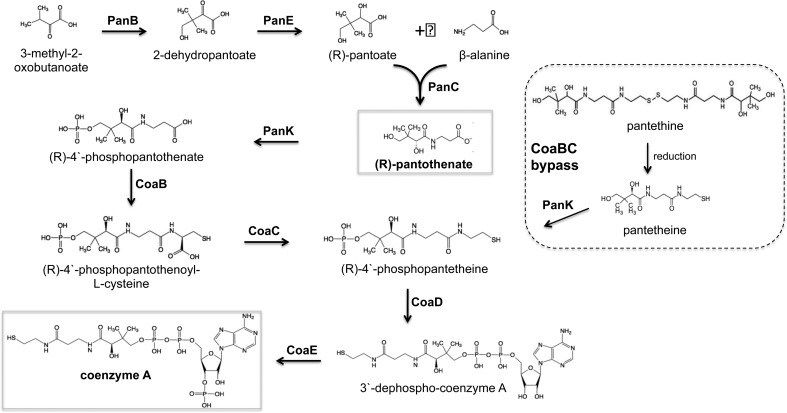
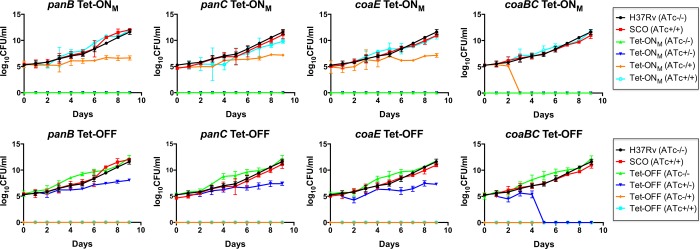
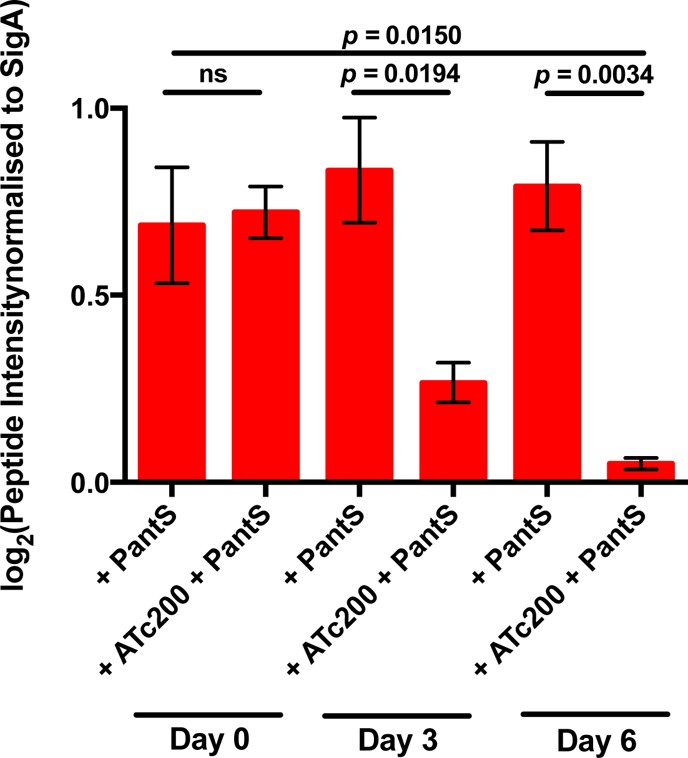
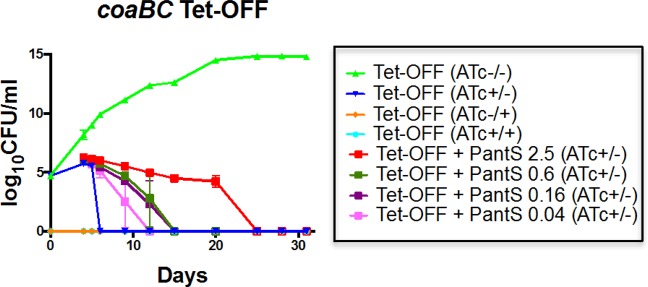
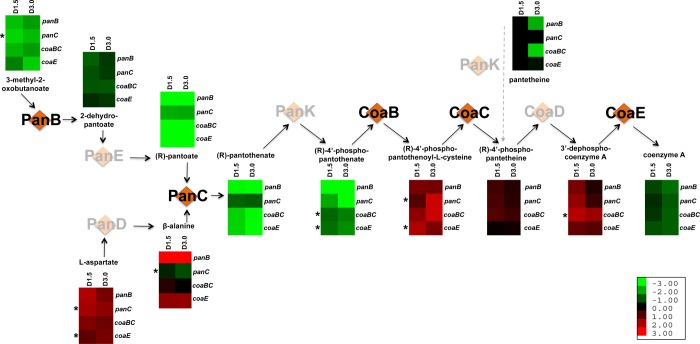
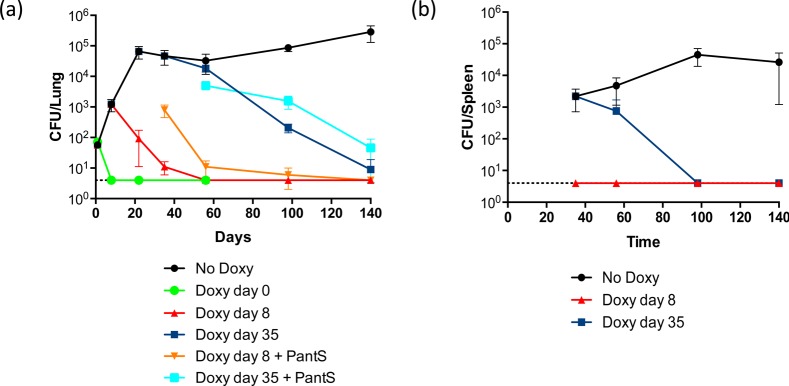
Mycobacterium tuberculosis relies on its own ability to biosynthesize coenzyme A to meet the needs of the myriad enzymatic reactions that depend on this cofactor for activity. As such, the essential pantothenate and coenzyme A biosynthesis pathways have attracted attention as targets for tuberculosis drug development. To identify the optimal step for coenzyme A pathway disruption in M. tuberculosis, we constructed and characterized a panel of conditional knockdown mutants in coenzyme A pathway genes. Here, we report that silencing of coaBC was bactericidal in vitro, whereas silencing of panB, panC, or coaE was bacteriostatic over the same time course. Silencing of coaBC was likewise bactericidal in vivo, whether initiated at infection or during either the acute or chronic stages of infection, confirming that CoaBC is required for M. tuberculosis to grow and persist in mice and arguing against significant CoaBC bypass via transport and assimilation of host-derived pantetheine in this animal model. These results provide convincing genetic validation of CoaBC as a new bactericidal drug target.
Keywords: CoA; CoaBC; drug discovery; pantetheine; tuberculosis.
Conflict of interest statement
The authors declare no competing financial interest.
Figures

References
-
- (2015) Global Tuberculosis Report, WHO.
-
- Andries K.; Verhasselt P.; Guillemont J.; Gohlmann H. W.; Neefs J. M.; Winkler H.; Van Gestel J.; Timmerman P.; Zhu M.; Lee E.; Williams P.; de Chaffoy D.; Huitric E.; Hoffner S.; Cambau E.; Truffot-Pernot C.; Lounis N.; Jarlier V. (2005) A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis. Science 307 (5707), 223–227. 10.1126/science.1106753. - DOI - PubMed
-
- Matsumoto M.; Hashizume H.; Tomishige T.; Kawasaki M.; Tsubouchi H.; Sasaki H.; Shimokawa Y.; Komatsu M. (2006) OPC-67683, a nitro-dihydro-imidazooxazole derivative with promising action against tuberculosis in vitro and in mice. PLoS Med. 3 (11), e466. 10.1371/journal.pmed.0030466. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
