

Ethylene-Regulated Glutamate Dehydrogenase Fine-Tunes Metabolism during Anoxia-Reoxygenation
- PMID: 27677986
- PMCID: PMC5100772
- DOI: 10.1104/pp.16.00985
Ethylene-Regulated Glutamate Dehydrogenase Fine-Tunes Metabolism during Anoxia-Reoxygenation
Abstract
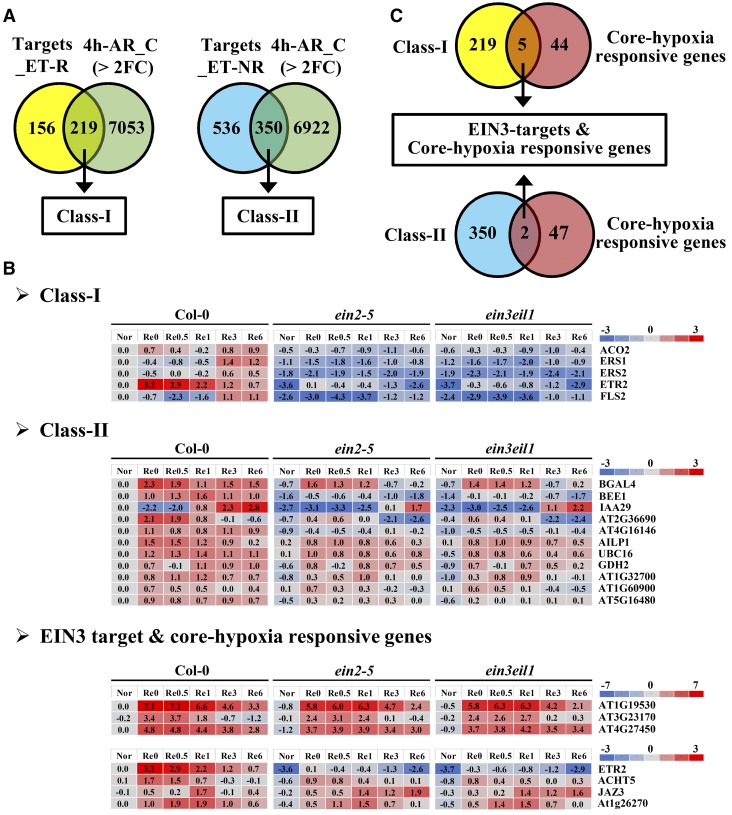
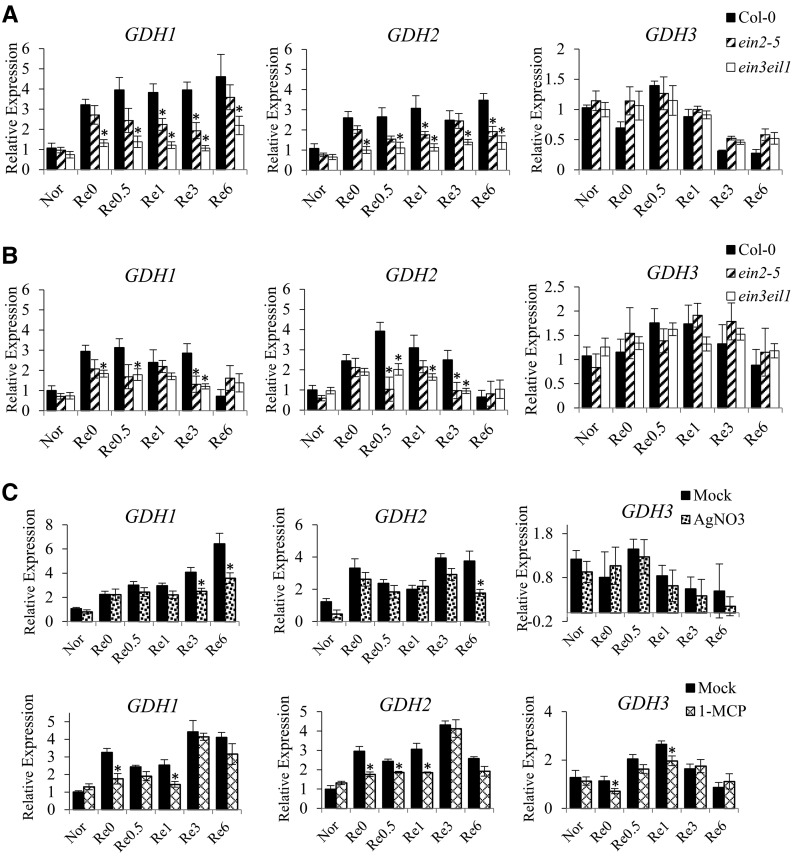
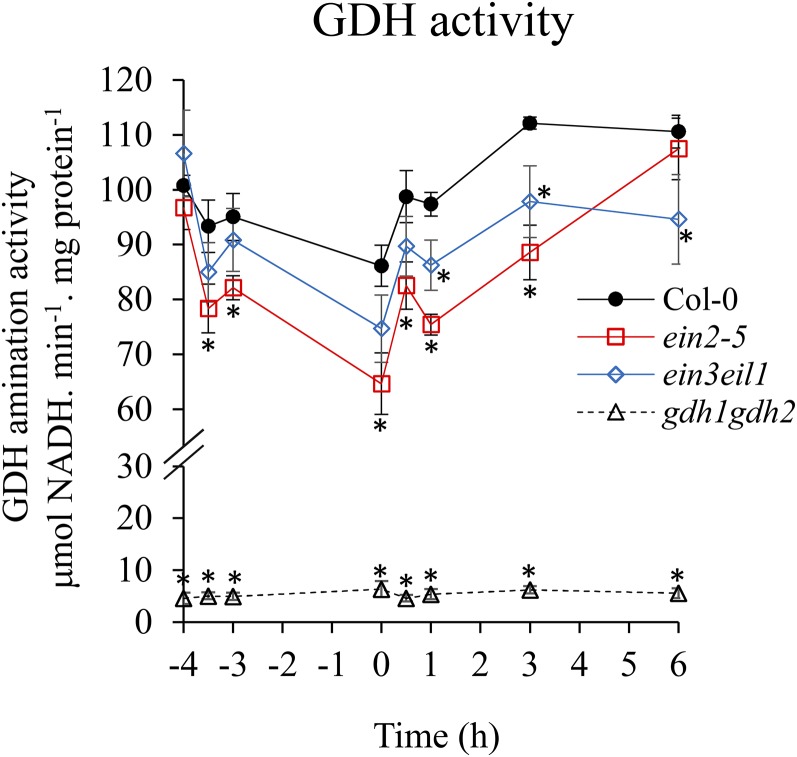
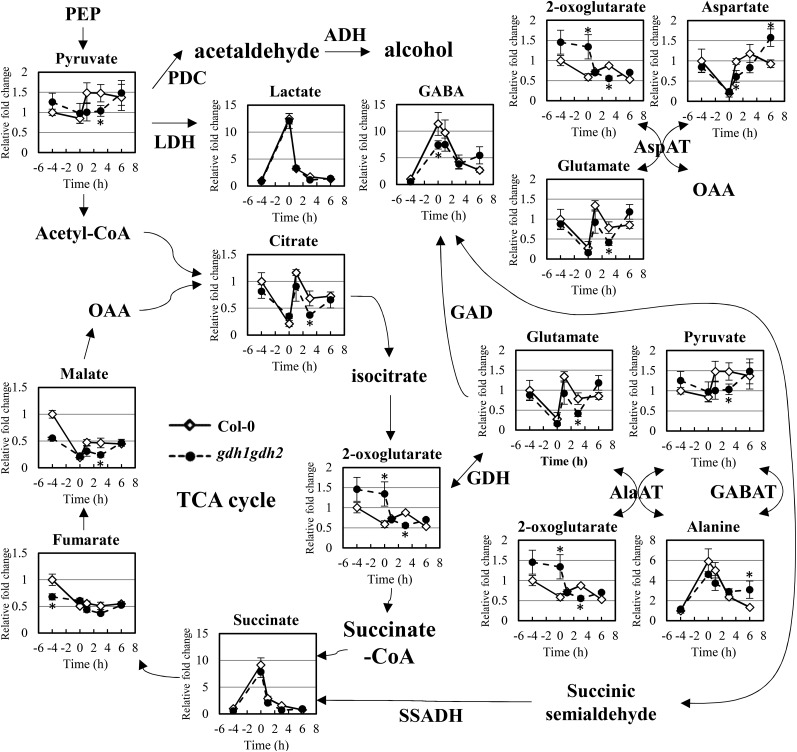
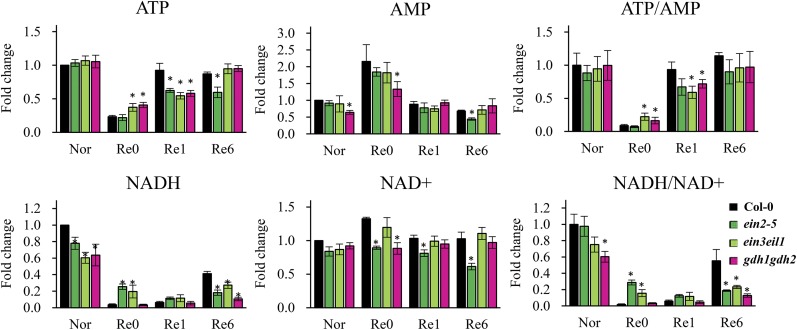
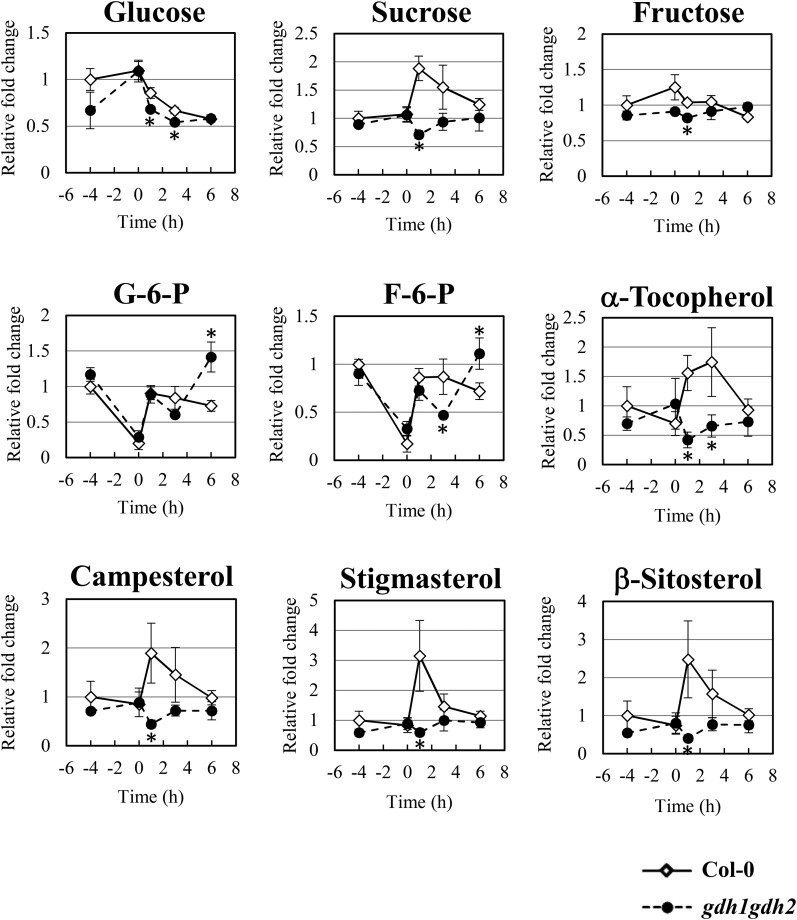
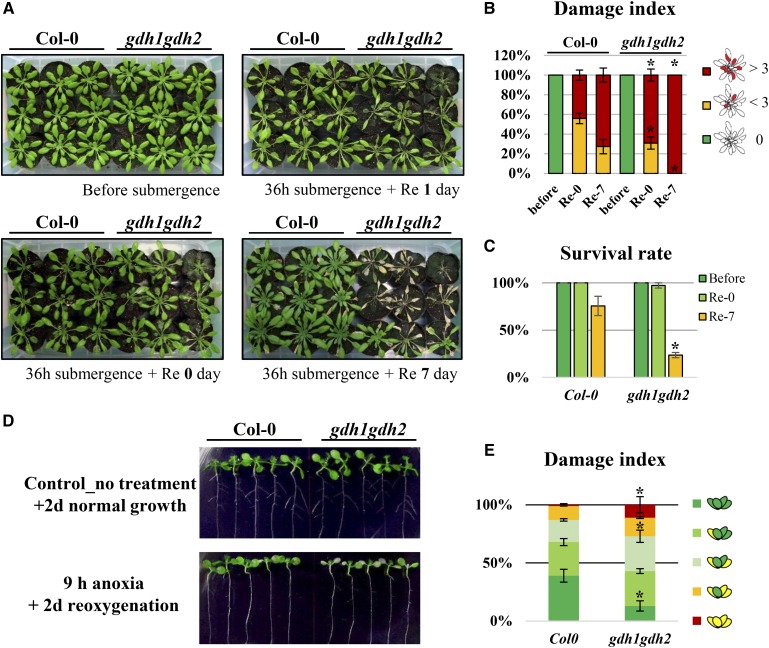
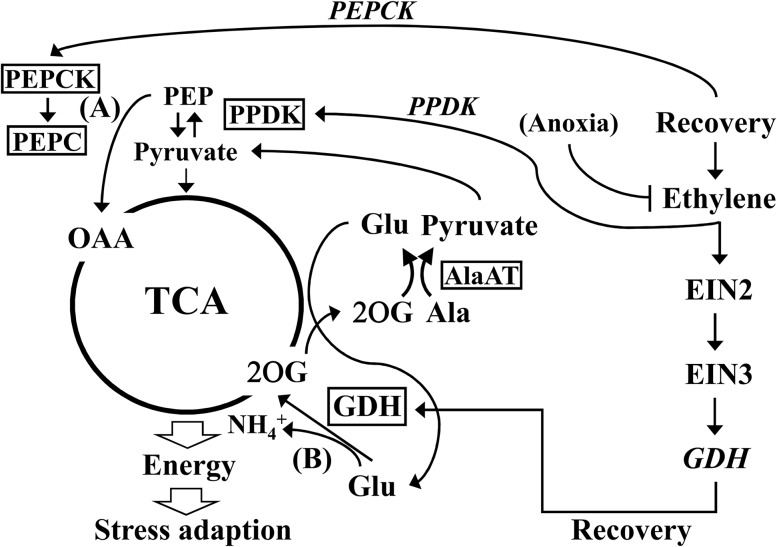
Ethylene is an essential hormone in plants that is involved in low-oxygen and reoxygenation responses. As a key transcription factor in ethylene signaling, ETHYLENE INSENSITIVE3 (EIN3) activates targets that trigger various responses. However, most of these targets are still poorly characterized. Through analyses of our microarray data and the published Arabidopsis (Arabidopsis thaliana) EIN3 chromatin immunoprecipitation sequencing data set, we inferred the putative targets of EIN3 during anoxia-reoxygenation. Among them, GDH2, which encodes one subunit of glutamate dehydrogenase (GDH), was chosen for further studies for its role in tricarboxylic acid cycle replenishment. We demonstrated that both GDH1 and GDH2 are induced during anoxia and reoxygenation and that this induction is mediated via ethylene signaling. In addition, the results of enzymatic assays showed that the level of GDH during anoxia-reoxygenation decreased in the ethylene-insensitive mutants ein2-5 and ein3eil1 Global metabolite analysis indicated that the deamination activity of GDH might regenerate 2-oxoglutarate, which is a cosubstrate that facilitates the breakdown of alanine by alanine aminotransferase when reoxygenation occurs. Moreover, ineffective tricarboxylic acid cycle replenishment, disturbed carbohydrate metabolism, reduced phytosterol biosynthesis, and delayed energy regeneration were found in gdh1gdh2 and ethylene mutants during reoxygenation. Taken together, these data illustrate the essential role of EIN3-regulated GDH activity in metabolic adjustment during anoxia-reoxygenation.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures

References
-
- Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR (1999) EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284: 2148–2152 - PubMed
-
- Bieniawska Z, Barratt DHP, Garlick AP, Thole V, Kruger NJ, Martin C, Zrenner R, Smith AM (2007) Analysis of the sucrose synthase gene family in Arabidopsis. Plant J 49: 810–828 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
