

Intracellular mechanisms of molecular recognition and sorting for transport of large extracellular matrix molecules
- PMID: 27679847
- PMCID: PMC5068301
- DOI: 10.1073/pnas.1609571113
Intracellular mechanisms of molecular recognition and sorting for transport of large extracellular matrix molecules
Abstract
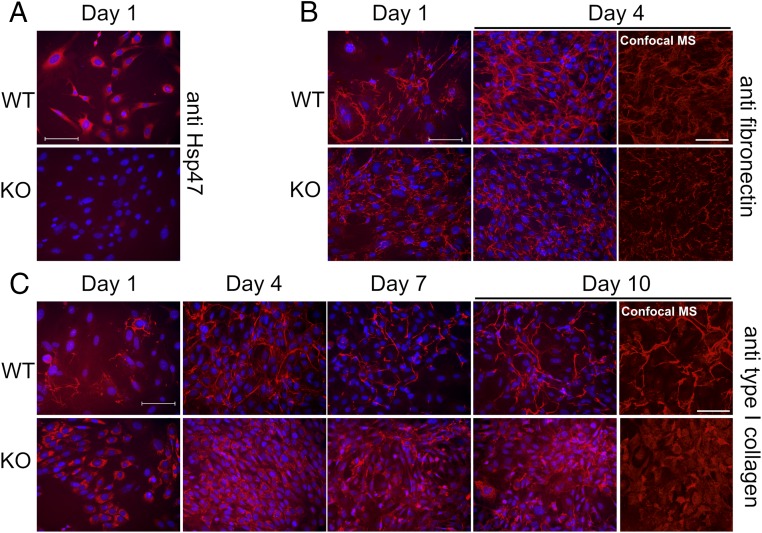
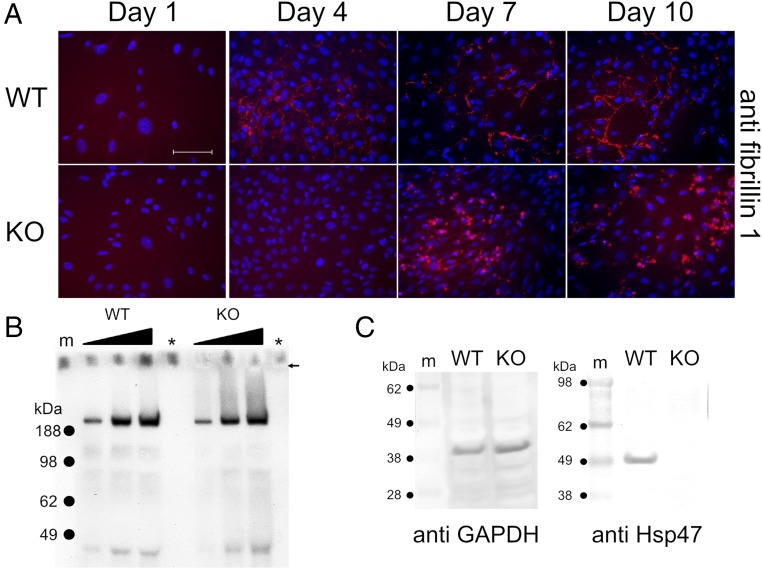
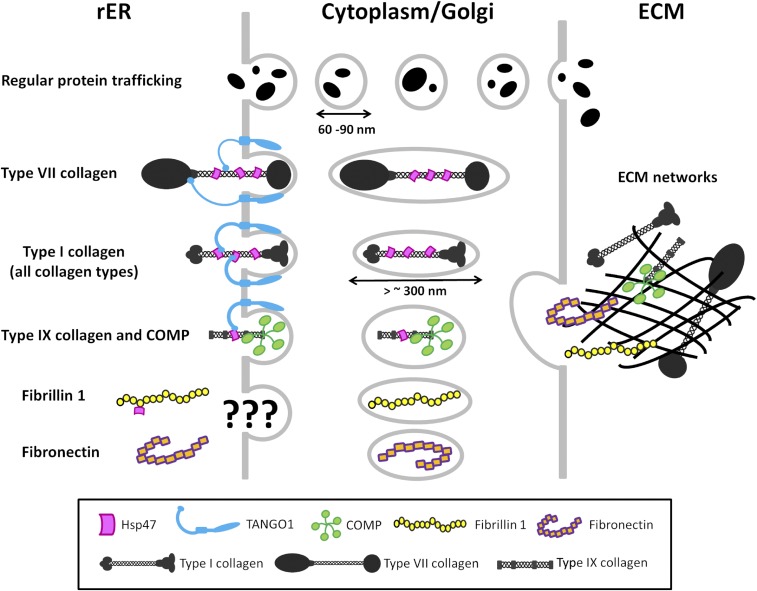
Extracellular matrix (ECM) proteins are biosynthesized in the rough endoplasmic reticulum (rER) and transported via the Golgi apparatus to the extracellular space. The coat protein complex II (COPII) transport vesicles are approximately 60-90 nm in diameter. However, several ECM molecules are much larger, up to several hundreds of nanometers. Therefore, special COPII vesicles are required to coat and transport these molecules. Transmembrane Protein Transport and Golgi Organization 1 (TANGO1) facilitates loading of collagens into special vesicles. The Src homology 3 (SH3) domain of TANGO1 was proposed to recognize collagen molecules, but how the SH3 domain recognizes various types of collagen is not understood. Moreover, how are large noncollagenous ECM molecules transported from the rER to the Golgi? Here we identify heat shock protein (Hsp) 47 as a guide molecule directing collagens to special vesicles by interacting with the SH3 domain of TANGO1. We also consider whether the collagen secretory model applies to other large ECM molecules.
Keywords: COPII vesicles; Hsp47; TANGO1; collagen; secretion.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
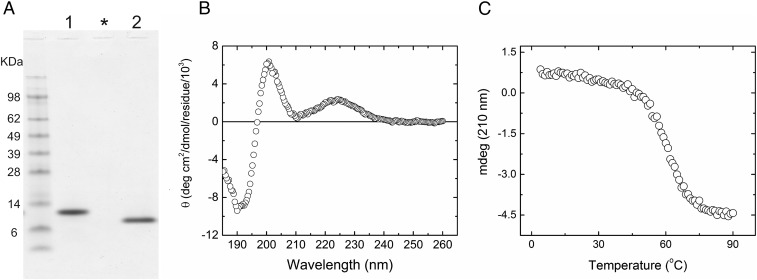
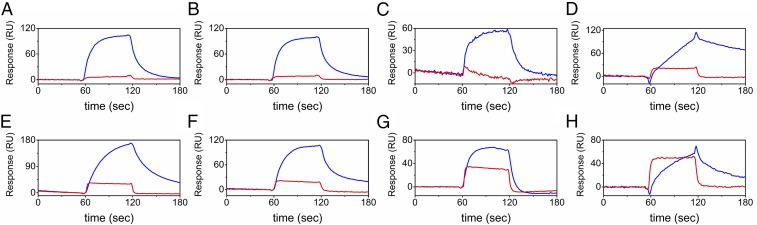
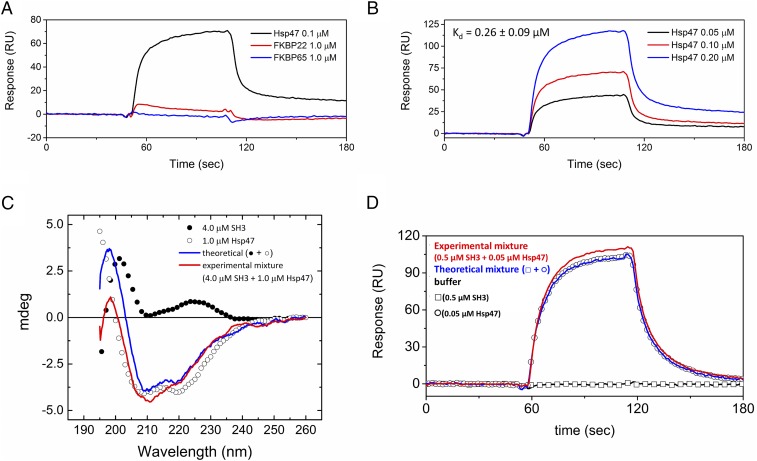
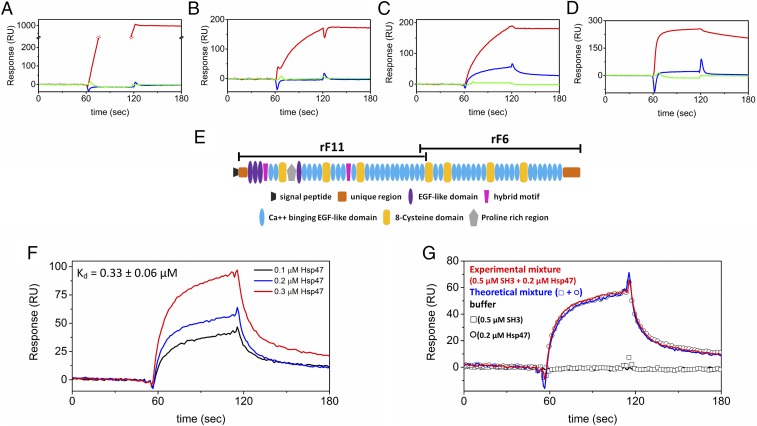
References
-
- Bateman JF, Boot-Handford RP, Lamandé SR. Genetic diseases of connective tissues: Cellular and extracellular effects of ECM mutations. Nat Rev Genet. 2009;10(3):173–183. - PubMed
-
- Bächinger HP, Mizuno K, Vranka J, Boudko S. Collagen formation and structure. In: Mander L, Liu HW, editors. Comprehensive Natural Products II: Chemistry and Biology. Elsevier; Amsterdam: 2010. pp. 469–530.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
