

Loss of ift122, a Retrograde Intraflagellar Transport (IFT) Complex Component, Leads to Slow, Progressive Photoreceptor Degeneration Due to Inefficient Opsin Transport
- PMID: 27681595
- PMCID: PMC5114401
- DOI: 10.1074/jbc.M116.738658
Loss of ift122, a Retrograde Intraflagellar Transport (IFT) Complex Component, Leads to Slow, Progressive Photoreceptor Degeneration Due to Inefficient Opsin Transport
Abstract
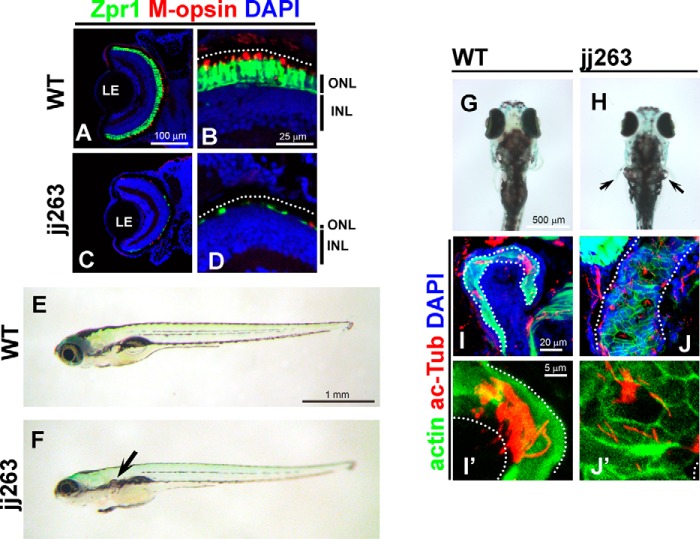
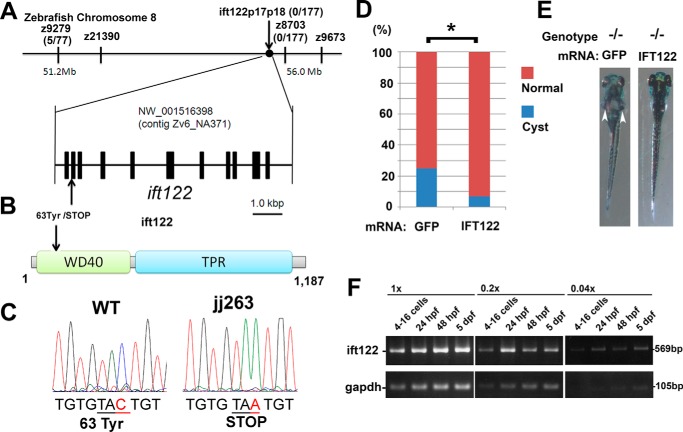
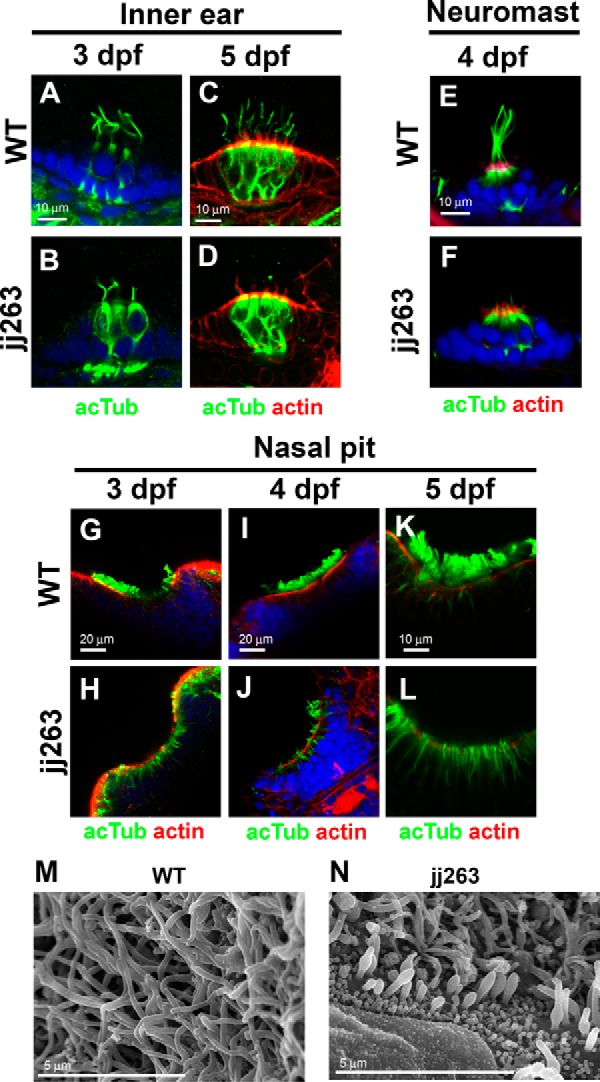
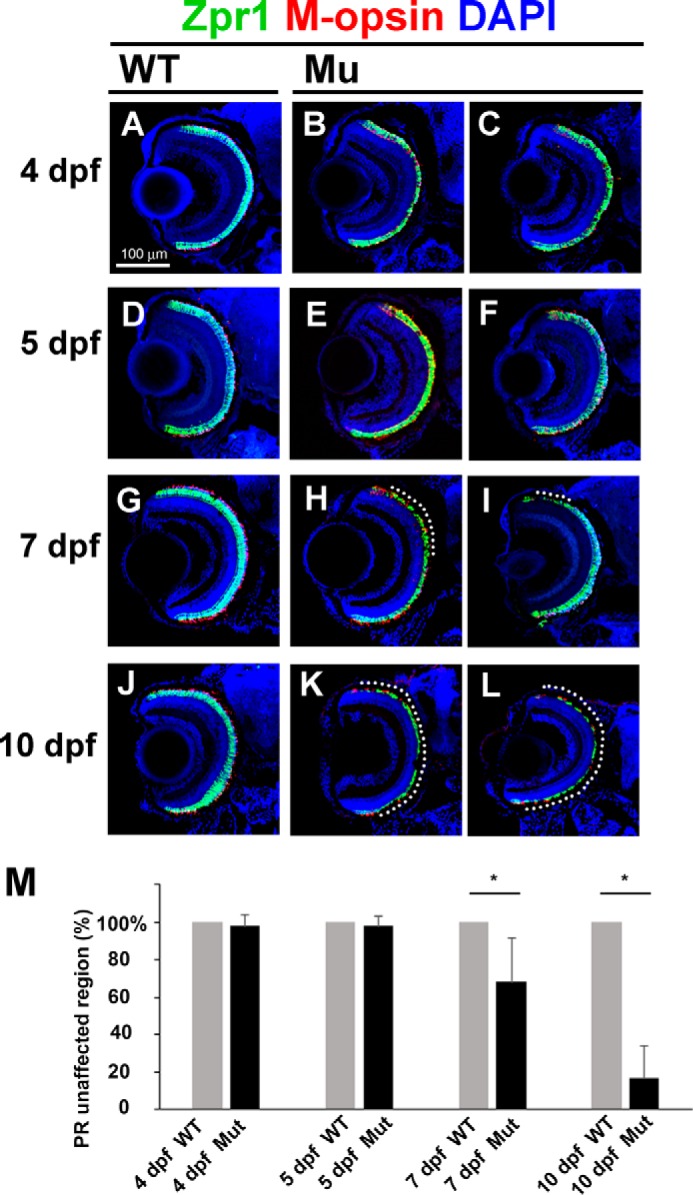
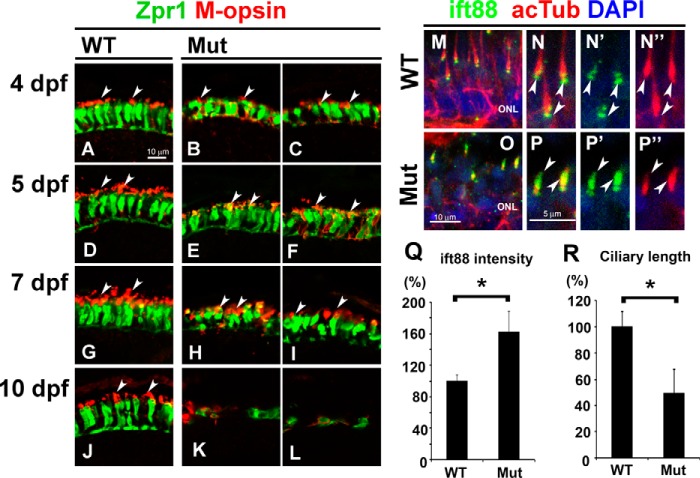
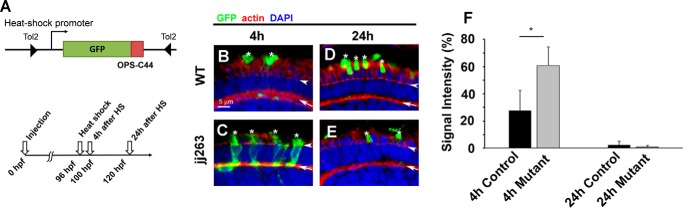
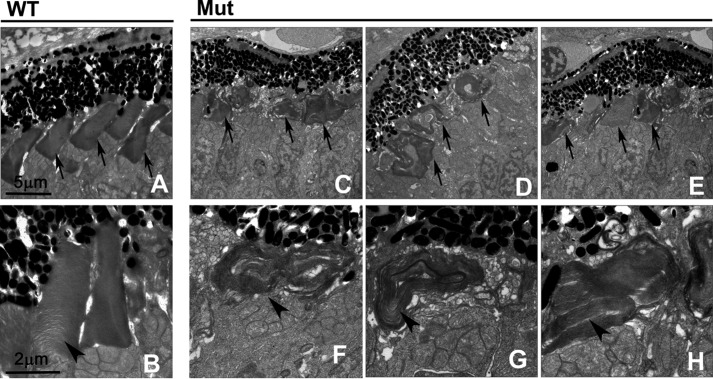
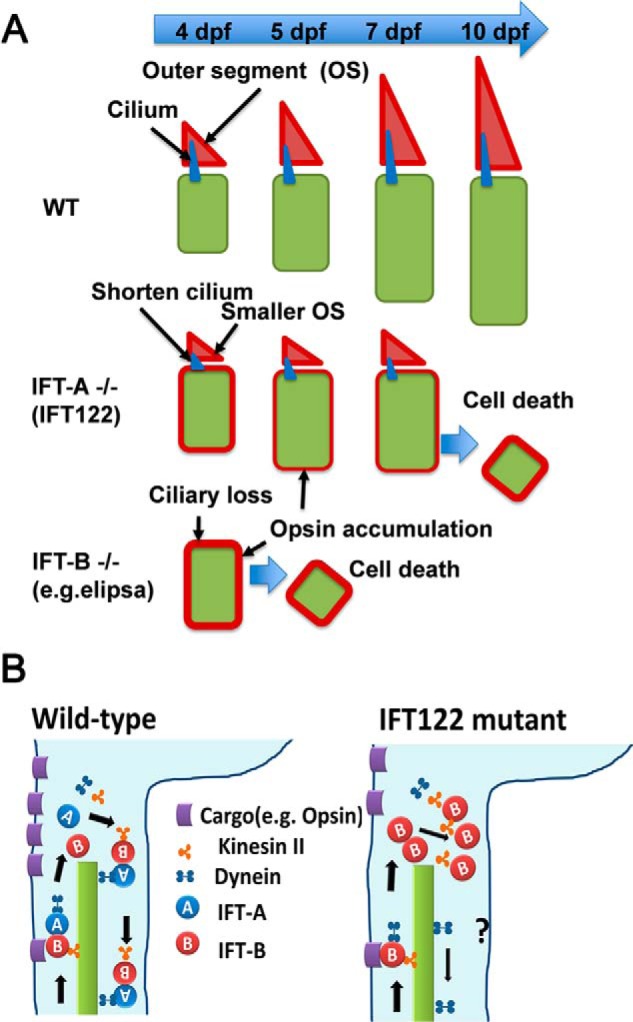
In the retina, aberrant opsin transport from cell bodies to outer segments leads to retinal degenerative diseases such as retinitis pigmentosa. Opsin transport is facilitated by the intraflagellar transport (IFT) system that mediates the bidirectional movement of proteins within cilia. In contrast to functions of the anterograde transport executed by IFT complex B (IFT-B), the precise functions of the retrograde transport mediated by IFT complex A (IFT-A) have not been well studied in photoreceptor cilia. Here, we analyzed developing zebrafish larvae carrying a null mutation in ift122 encoding a component of IFT-A. ift122 mutant larvae show unexpectedly mild phenotypes, compared with those of mutants defective in IFT-B. ift122 mutants exhibit a slow onset of progressive photoreceptor degeneration mainly after 7 days post-fertilization. ift122 mutant larvae also develop cystic kidney but not curly body, both of which are typically observed in various ciliary mutants. ift122 mutants display a loss of cilia in the inner ear hair cells and nasal pit epithelia. Loss of ift122 causes disorganization of outer segment discs. Ectopic accumulation of an IFT-B component, ift88, is observed in the ift122 mutant photoreceptor cilia. In addition, pulse-chase experiments using GFP-opsin fusion proteins revealed that ift122 is required for the efficient transport of opsin and the distal elongation of outer segments. These results show that IFT-A is essential for the efficient transport of outer segment proteins, including opsin, and for the survival of retinal photoreceptor cells, rendering the ift122 mutant a unique model for human retinal degenerative diseases.
Keywords: cilia; hair cell; photoreceptor; retinal degeneration; rhodopsin.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Fliegauf M., Benzing T., and Omran H. (2007) When cilia go bad: cilia defects and ciliopathies. Nat. Rev. Mol. Cell Biol. 8, 880–893 - PubMed
-
- Nigg E. A., and Raff J. W. (2009) Centrioles, centrosomes, and cilia in health and disease. Cell 139, 663–678 - PubMed
-
- Rosenbaum J. L., and Witman G. B. (2002) Intraflagellar transport. Nat. Rev. Mol. Cell Biol. 3, 813–825 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
