

Regulation of divalent metal transporter-1 by serine phosphorylation
- PMID: 27681840
- PMCID: PMC5103878
- DOI: 10.1042/BCJ20160674
Regulation of divalent metal transporter-1 by serine phosphorylation
Abstract
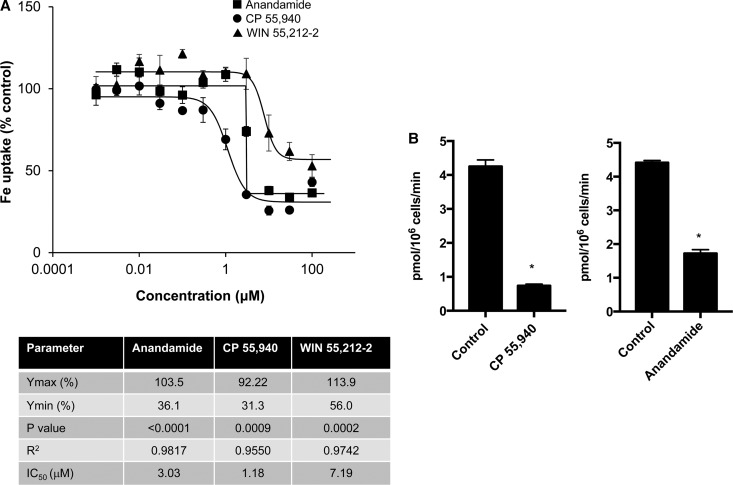
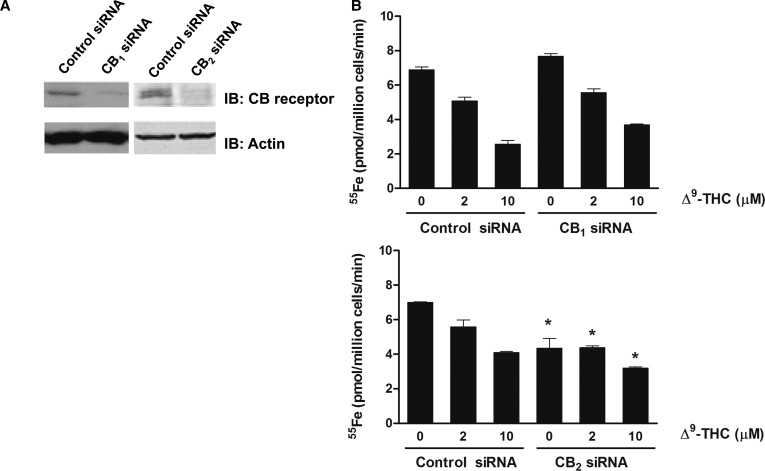
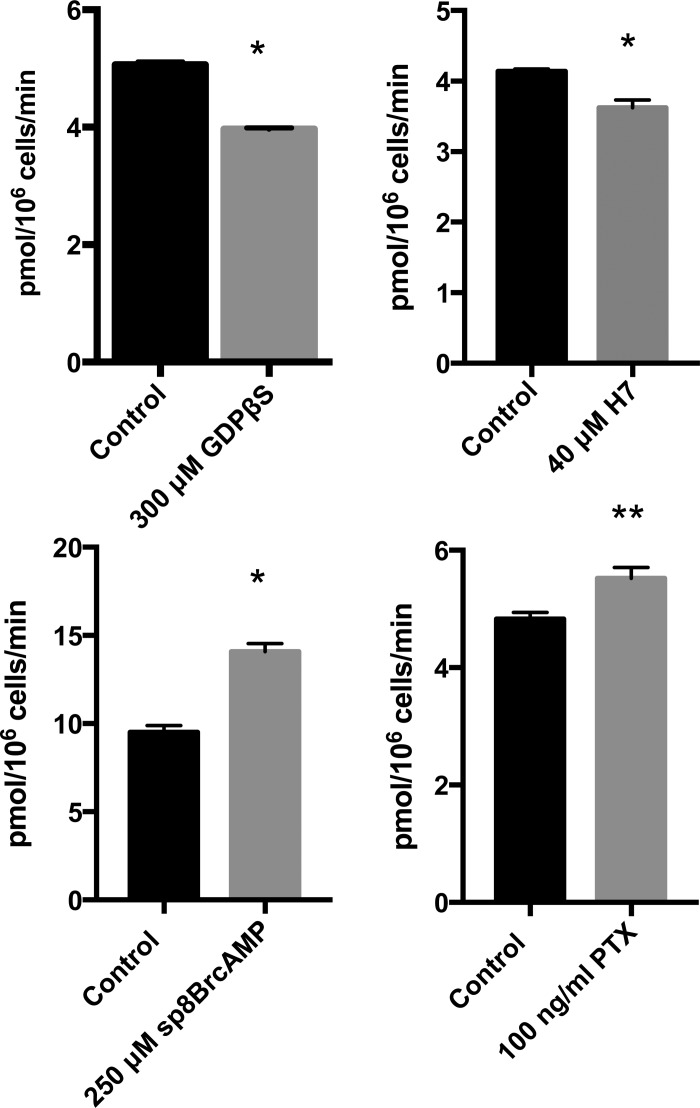
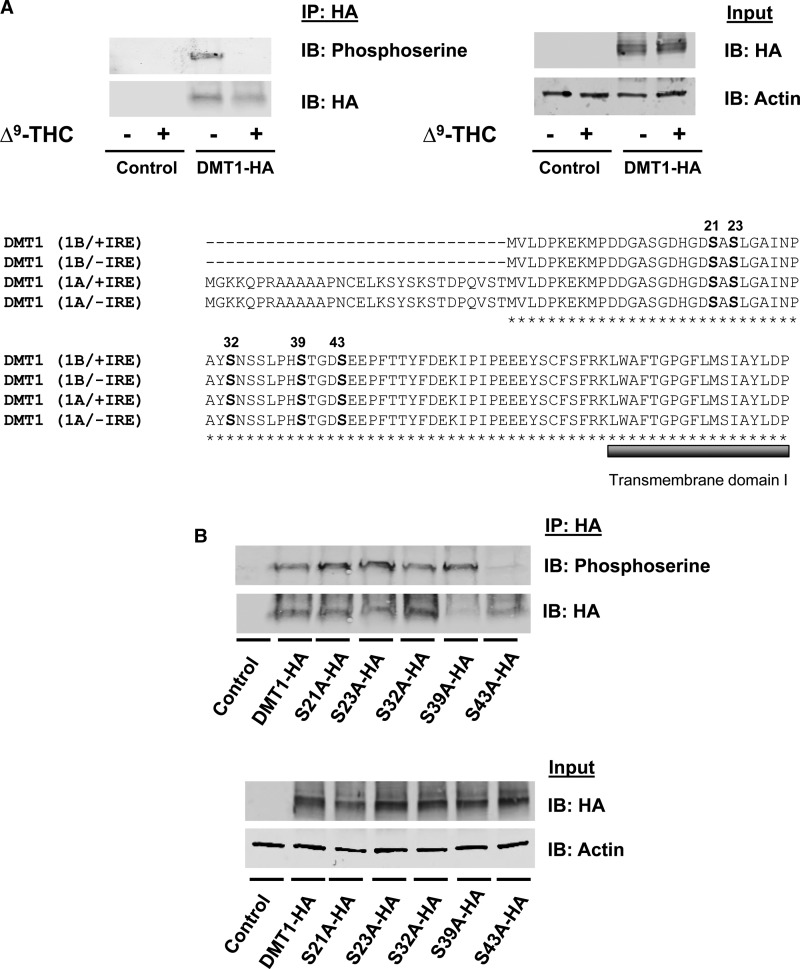
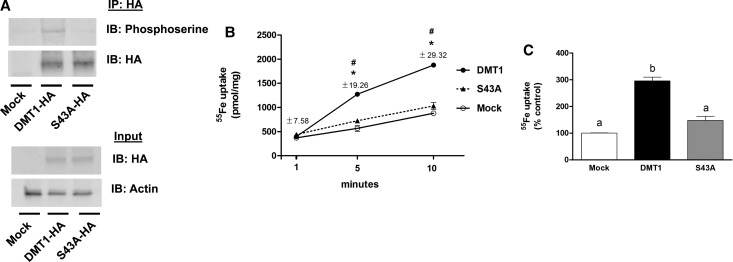
Divalent metal transporter-1 (DMT1) mediates dietary iron uptake across the intestinal mucosa and facilitates peripheral delivery of iron released by transferrin in the endosome. Here, we report that classical cannabinoids (Δ9-tetrahydrocannabinol, Δ9-THC), nonclassical cannabinoids (CP 55,940), aminoalkylindoles (WIN 55,212-2) and endocannabinoids (anandamide) reduce 55Fe and 54Mn uptake by HEK293T(DMT1) cells stably expressing the transporter. siRNA knockdown of cannabinoid receptor type 2 (CB2) abrogated inhibition. CB2 is a G-protein (GTP-binding protein)-coupled receptor that negatively regulates signal transduction cascades involving serine/threonine kinases. Immunoprecipitation experiments showed that DMT1 is serine-phosphorylated under basal conditions, but that treatment with Δ9-THC reduced phosphorylation. Site-directed mutation of predicted DMT1 phosphosites further showed that substitution of serine with alanine at N-terminal position 43 (S43A) abolished basal phosphorylation. Concordantly, both the rate and extent of 55Fe uptake in cells expressing DMT1(S43A) was reduced compared with those expressing wild-type DMT1. Among kinase inhibitors that affected DMT1-mediated iron uptake, staurosporine also reduced DMT1 phosphorylation confirming a role for serine phosphorylation in iron transport regulation. These combined data indicate that phosphorylation at serine 43 of DMT1 promotes transport activity, whereas dephosphorylation is associated with loss of iron uptake. Since anti-inflammatory actions mediated through CB2 would be associated with reduced DMT1 phosphorylation, we postulate that this pathway provides a means to reduce oxidative stress by limiting iron uptake.
Keywords: DMT1; Slc11a2; cannabinoid receptor; iron transport; Δ9-tetrahydrocannabinol.
© 2016 The Author(s).
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous