

Adipose Tissue Dendritic Cells Are Independent Contributors to Obesity-Induced Inflammation and Insulin Resistance
- PMID: 27683748
- PMCID: PMC5555636
- DOI: 10.4049/jimmunol.1600820
Adipose Tissue Dendritic Cells Are Independent Contributors to Obesity-Induced Inflammation and Insulin Resistance
Abstract
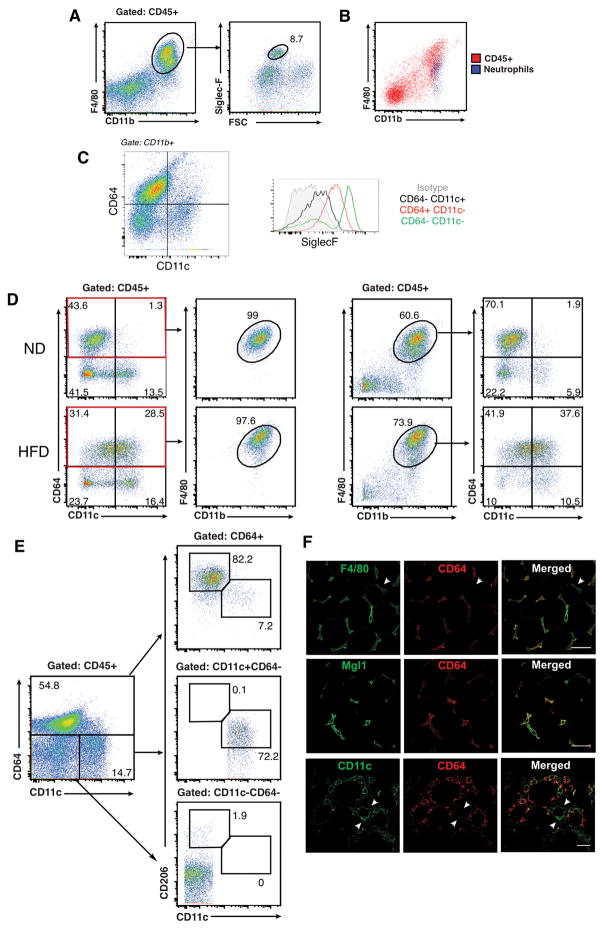
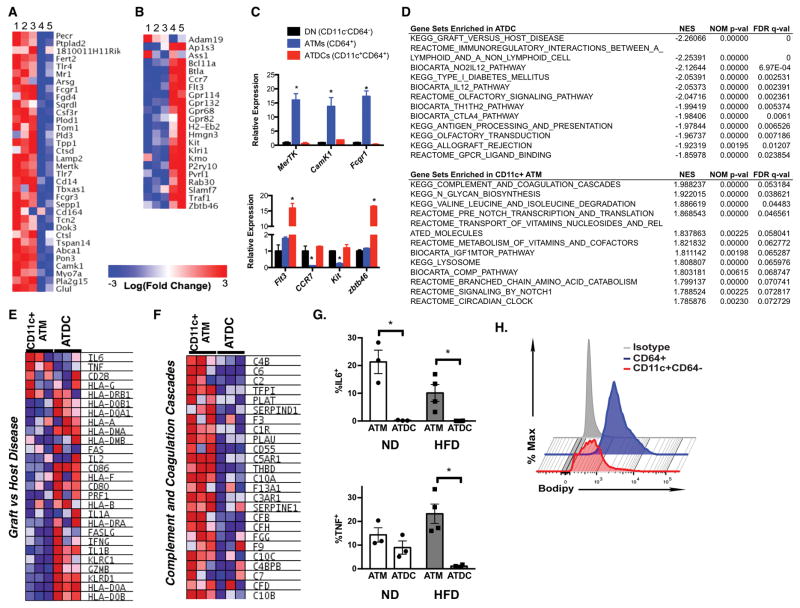
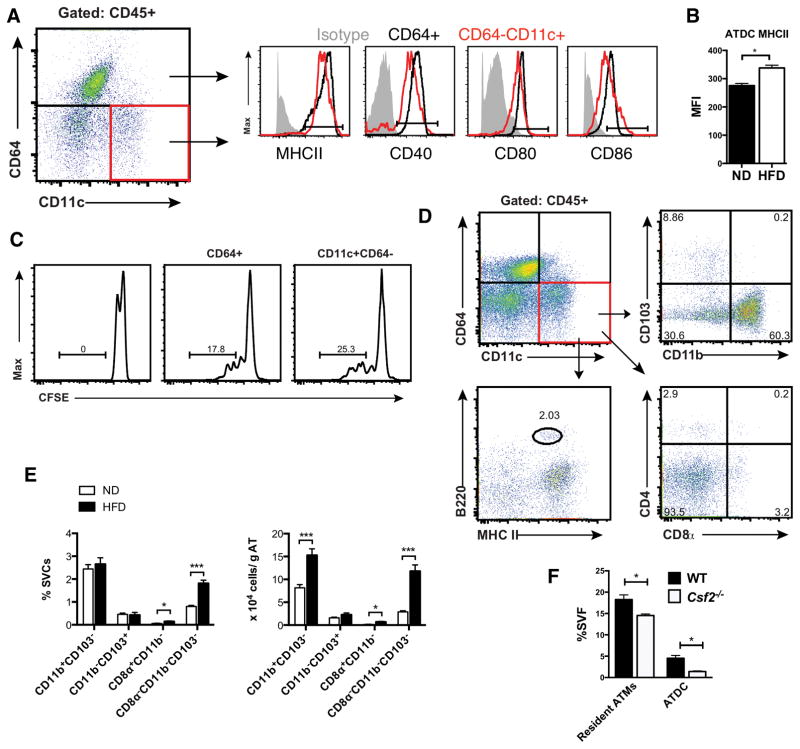
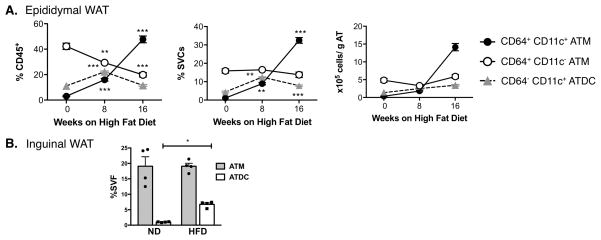
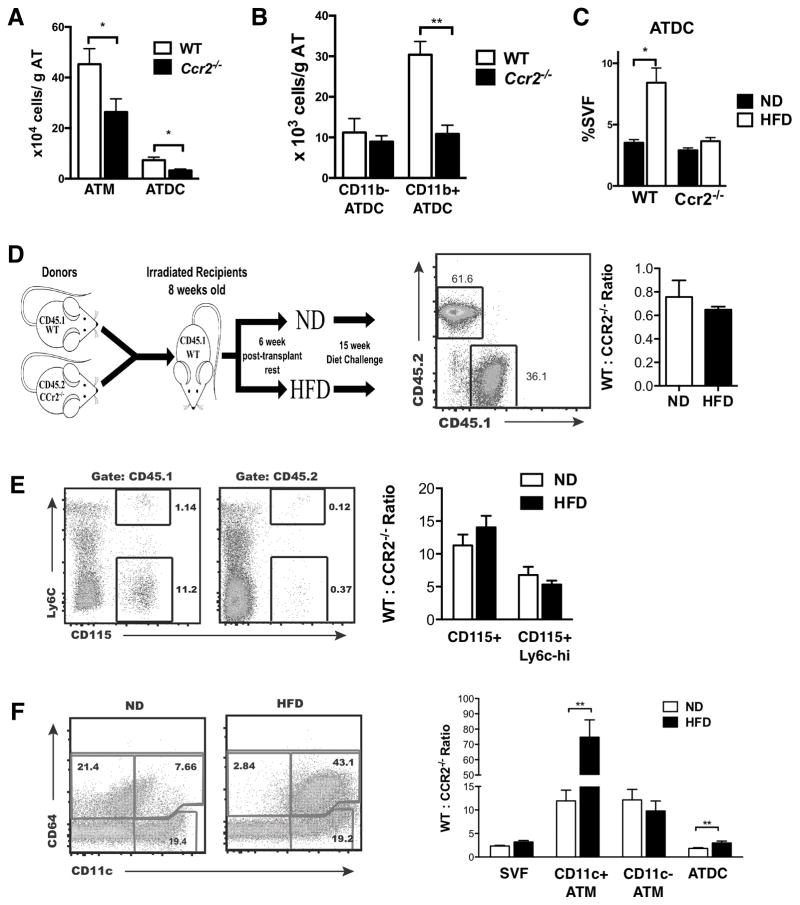
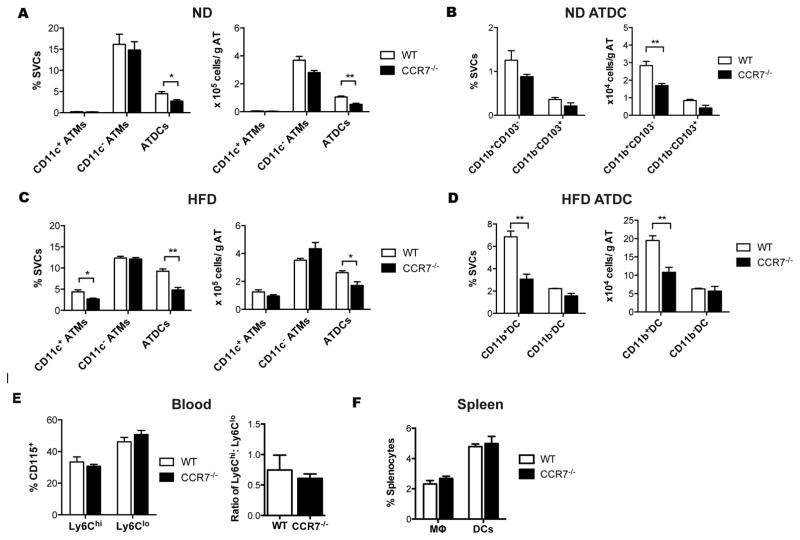
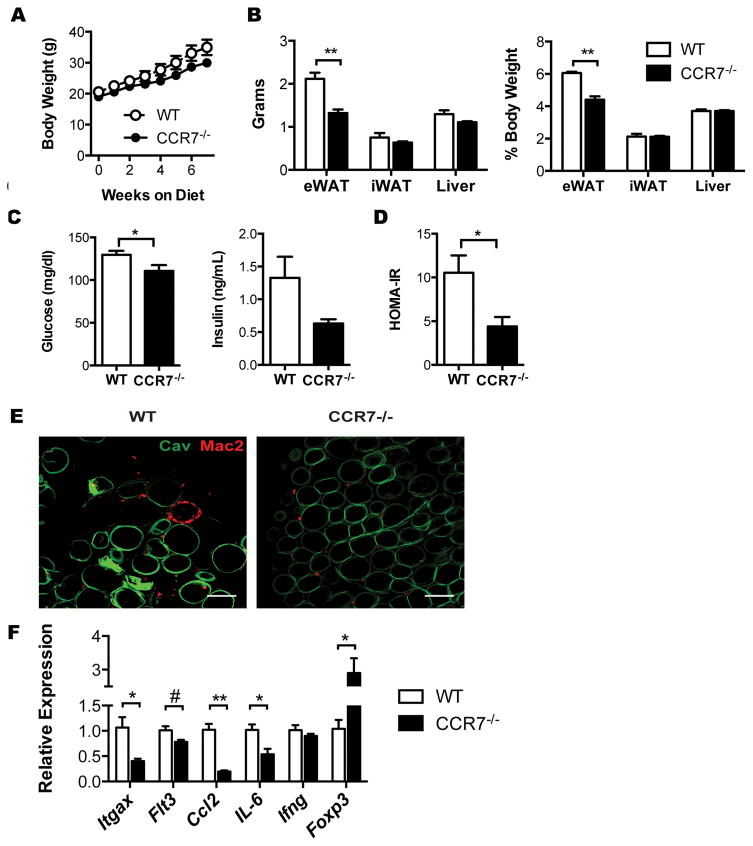
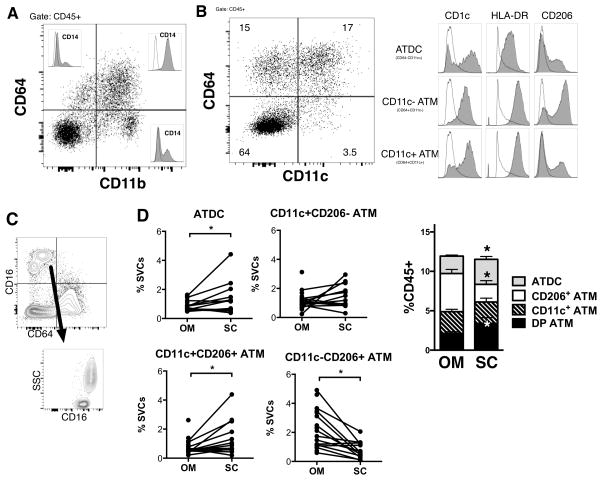
Dynamic changes of adipose tissue leukocytes, including adipose tissue macrophage (ATM) and adipose tissue dendritic cells (ATDCs), contribute to obesity-induced inflammation and metabolic disease. However, clear discrimination between ATDC and ATM in adipose tissue has limited progress in the field of immunometabolism. In this study, we use CD64 to distinguish ATM and ATDC, and investigated the temporal and functional changes in these myeloid populations during obesity. Flow cytometry and immunostaining demonstrated that the definition of ATM as F4/80+CD11b+ cells overlaps with other leukocytes and that CD45+CD64+ is specific for ATM. The expression of core dendritic cell genes was enriched in CD11c+CD64- cells (ATDC), whereas core macrophage genes were enriched in CD45+CD64+ cells (ATM). CD11c+CD64- ATDCs expressed MHC class II and costimulatory receptors, and had similar capacity to stimulate CD4+ T cell proliferation as ATMs. ATDCs were predominantly CD11b+ conventional dendritic cells and made up the bulk of CD11c+ cells in adipose tissue with moderate high-fat diet exposure. Mixed chimeric experiments with Ccr2-/- mice demonstrated that high-fat diet-induced ATM accumulation from monocytes was dependent on CCR2, whereas ATDC accumulation was less CCR2 dependent. ATDC accumulation during obesity was attenuated in Ccr7-/- mice and was associated with decreased adipose tissue inflammation and insulin resistance. CD45+CD64+ ATM and CD45+CD64-CD11c+ ATDCs were identified in human obese adipose tissue and ATDCs were increased in s.c. adipose tissue compared with omental adipose tissue. These results support a revised strategy for unambiguous delineation of ATM and ATDC, and suggest that ATDCs are independent contributors to adipose tissue inflammation during obesity.
Copyright © 2016 by The American Association of Immunologists, Inc.
Figures
References
-
- Poitou C, Dalmas E, Renovato M, Benhamo V, Hajduch F, Abdennour M, Kahn JF, Veyrie N, Rizkalla S, Fridman WH, Sautes-Fridman C, Clement K, Cremer I. CD14dimCD16+ and CD14+CD16+ monocytes in obesity and during weight loss: relationships with fat mass and subclinical atherosclerosis. Arterioscler Thromb Vac Biol. 2011;31:2322–2330. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
