

Auditory cortical field coding long-lasting tonal offsets in mice
- PMID: 27687766
- PMCID: PMC5043382
- DOI: 10.1038/srep34421
Auditory cortical field coding long-lasting tonal offsets in mice
Abstract
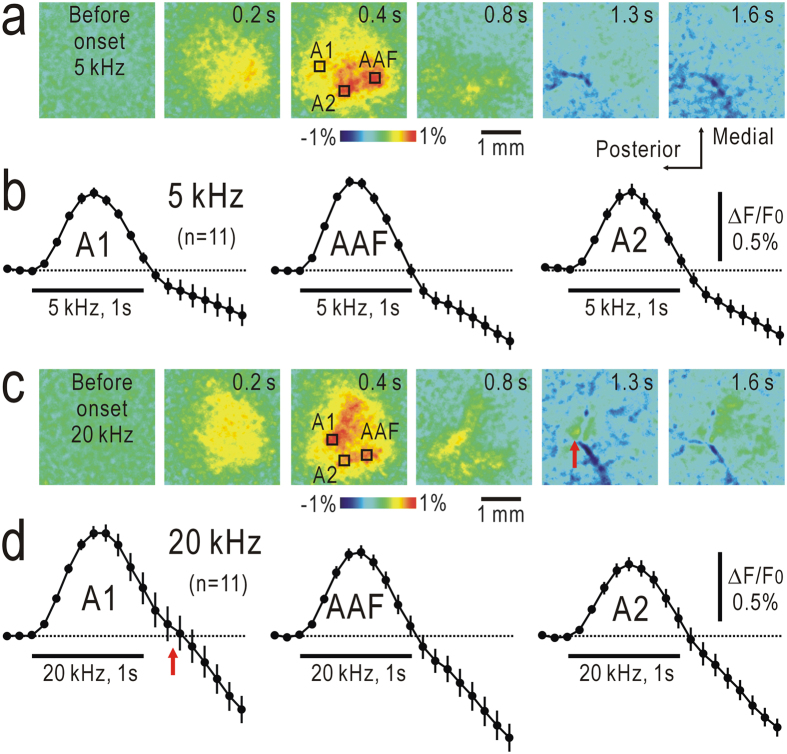
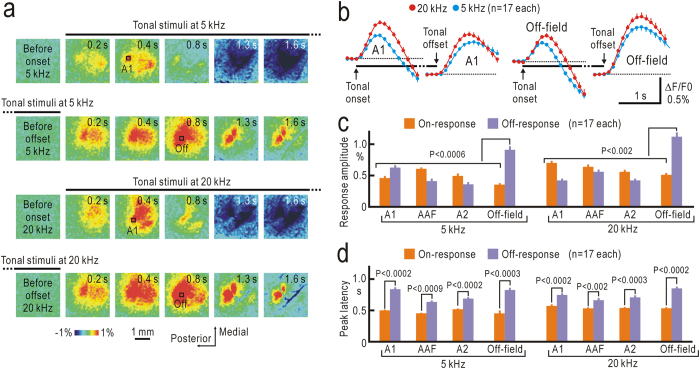
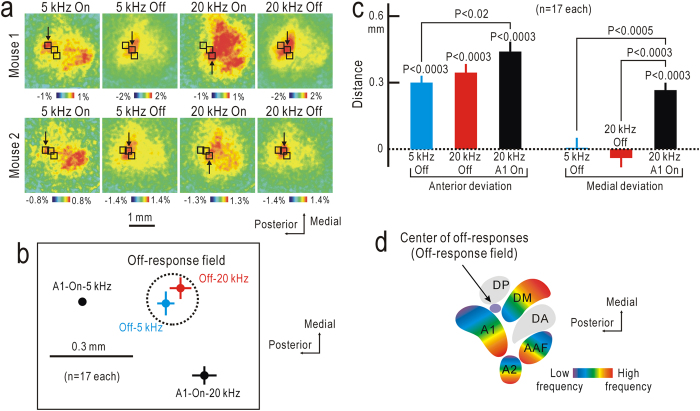
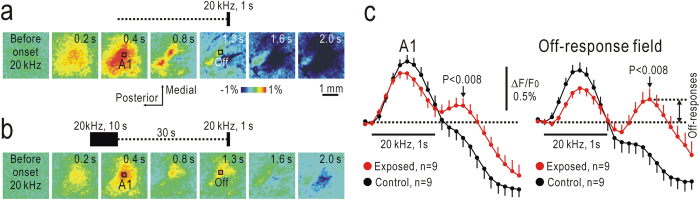
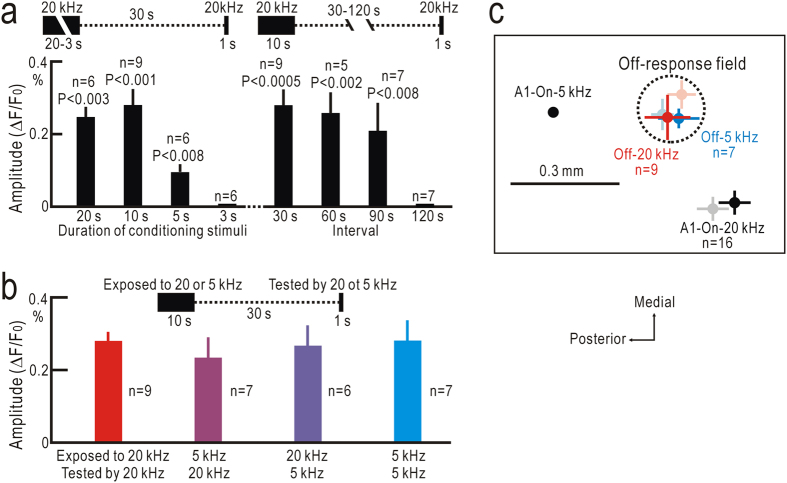
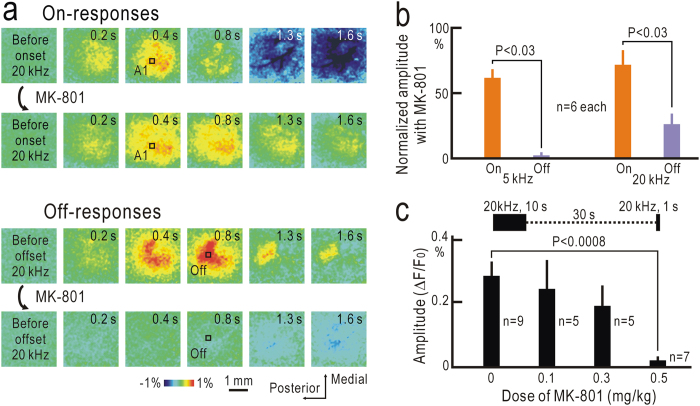
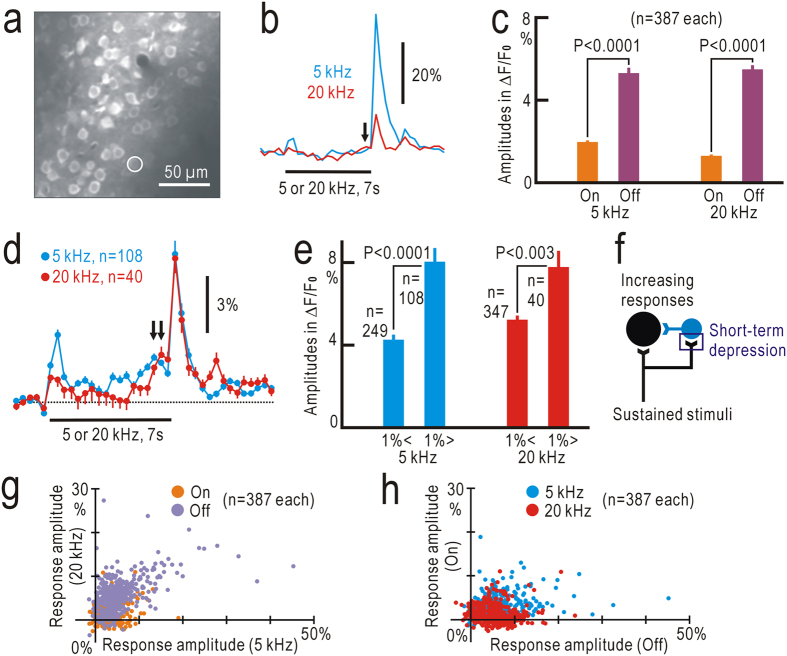
Although temporal information processing is important in auditory perception, the mechanisms for coding tonal offsets are unknown. We investigated cortical responses elicited at the offset of tonal stimuli using flavoprotein fluorescence imaging in mice. Off-responses were clearly observed at the offset of tonal stimuli lasting for 7 s, but not after stimuli lasting for 1 s. Off-responses to the short stimuli appeared in a similar cortical region, when conditioning tonal stimuli lasting for 5-20 s preceded the stimuli. MK-801, an inhibitor of NMDA receptors, suppressed the two types of off-responses, suggesting that disinhibition produced by NMDA receptor-dependent synaptic depression might be involved in the off-responses. The peak off-responses were localized in a small region adjacent to the primary auditory cortex, and no frequency-dependent shift of the response peaks was found. Frequency matching of preceding tonal stimuli with short test stimuli was not required for inducing off-responses to short stimuli. Two-photon calcium imaging demonstrated significantly larger neuronal off-responses to stimuli lasting for 7 s in this field, compared with off-responses to stimuli lasting for 1 s. The present results indicate the presence of an auditory cortical field responding to long-lasting tonal offsets, possibly for temporal information processing.
Figures
References
-
- Kudoh M., Seki K. & Shibuki K. Sound sequence discrimination learning is dependent on cholinergic inputs to the rat auditory cortex. Neurosci. Res. 50, 113–123 (2004). - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
