

Impact of Aging on Proprioceptive Sensory Neurons and Intrafusal Muscle Fibers in Mice
- PMID: 27688482
- PMCID: PMC6075202
- DOI: 10.1093/gerona/glw175
Impact of Aging on Proprioceptive Sensory Neurons and Intrafusal Muscle Fibers in Mice
Abstract
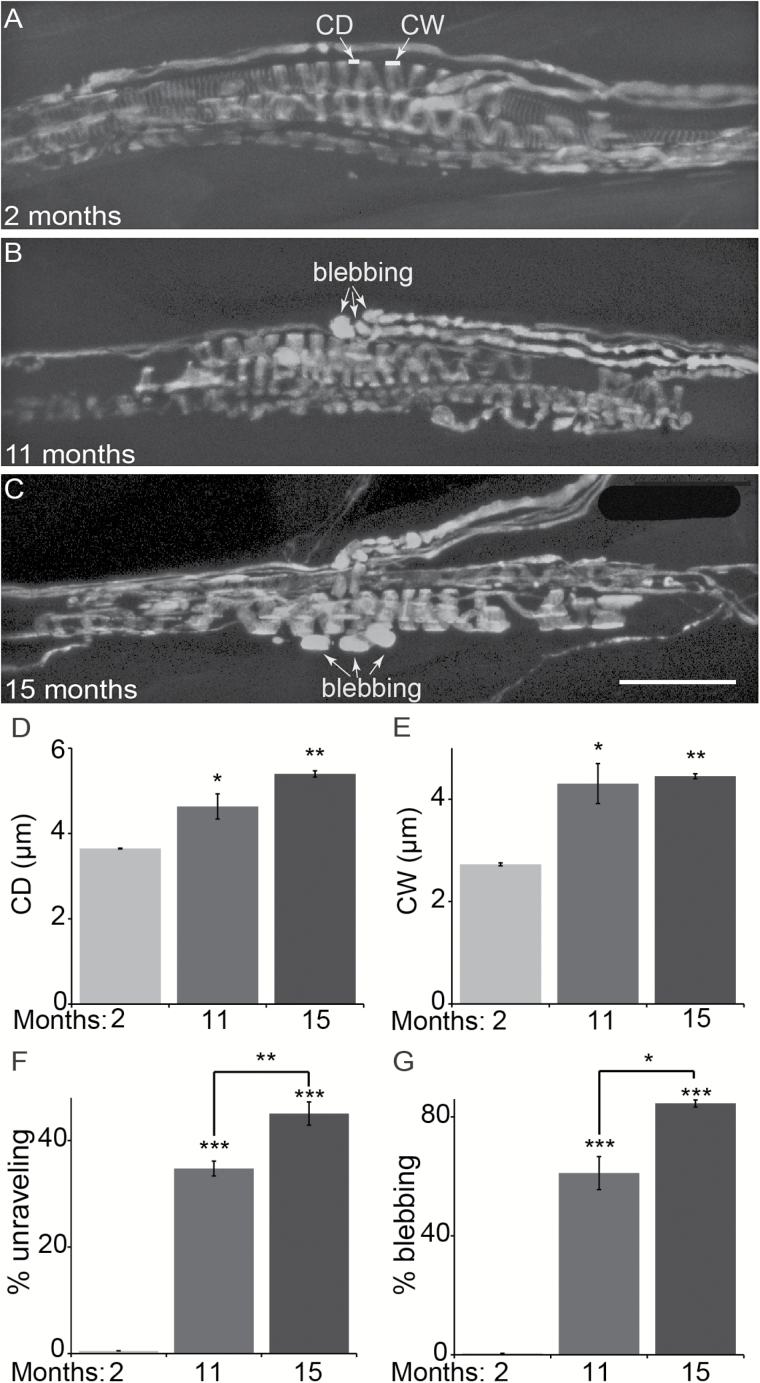
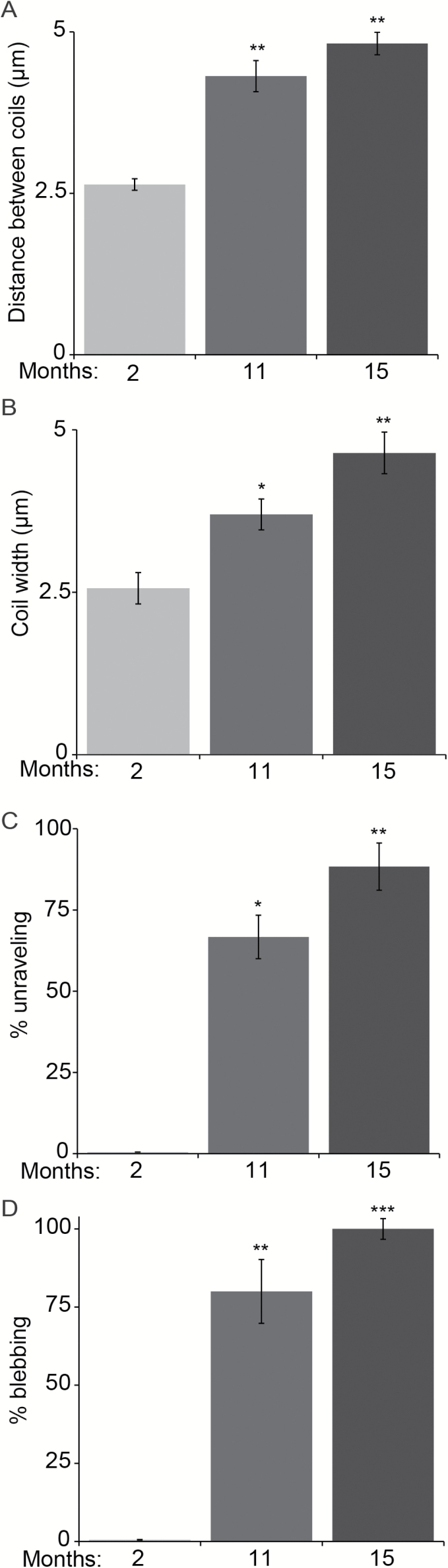
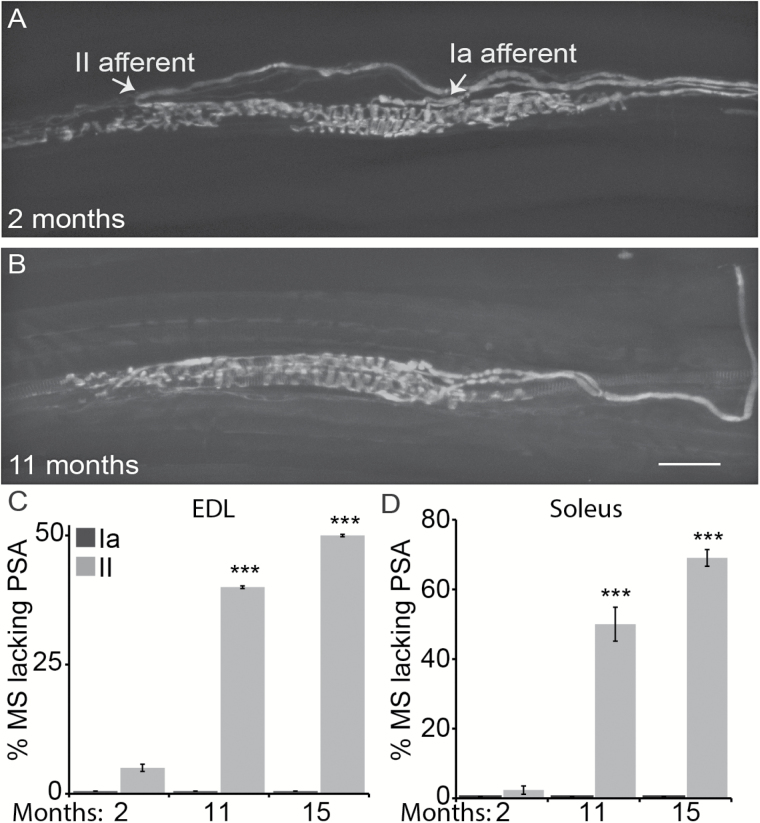
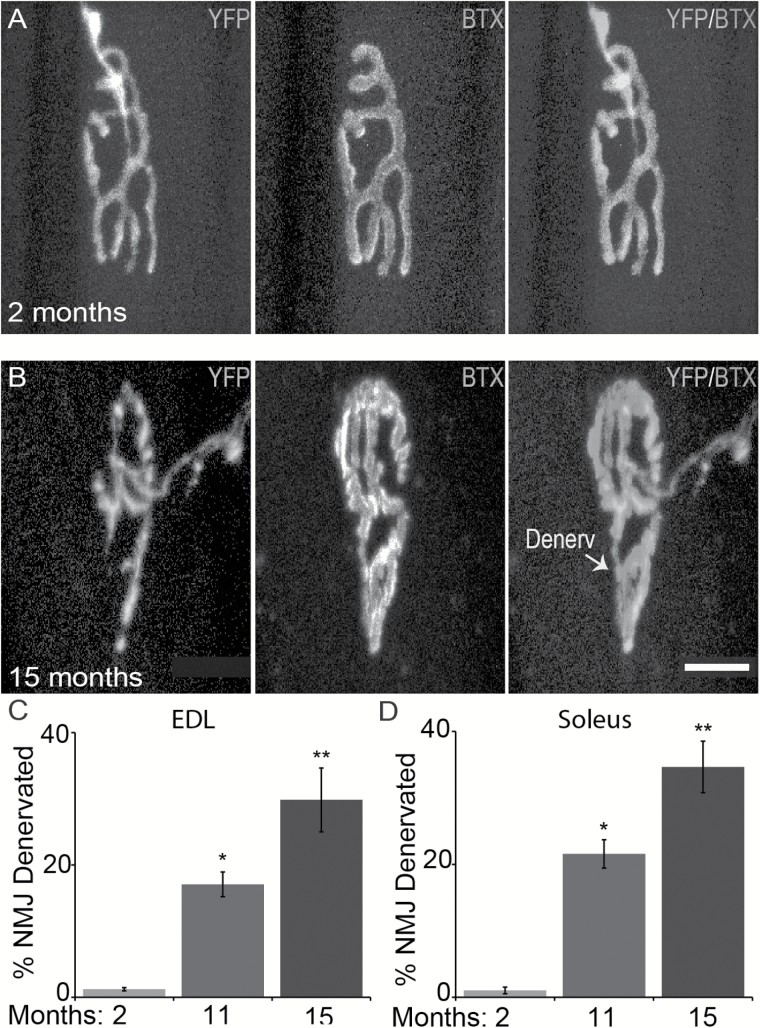
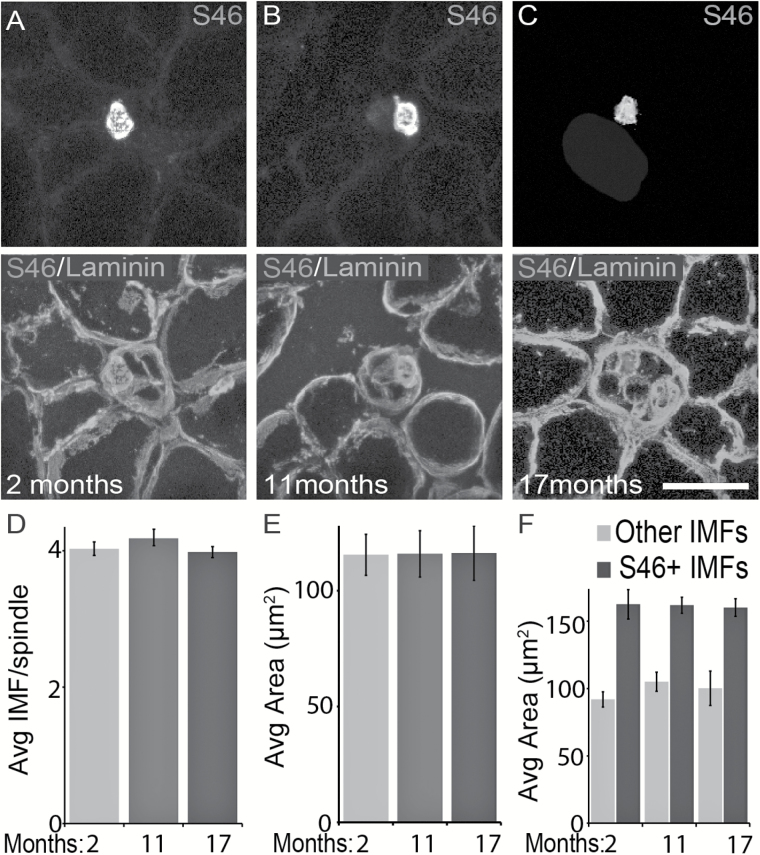
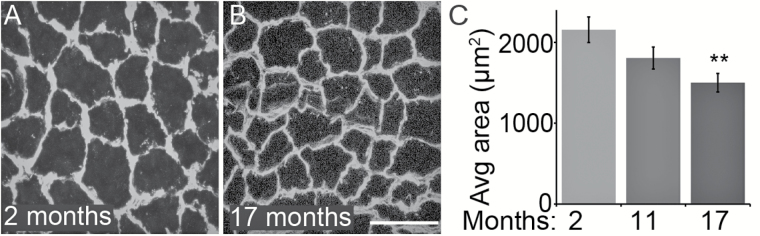
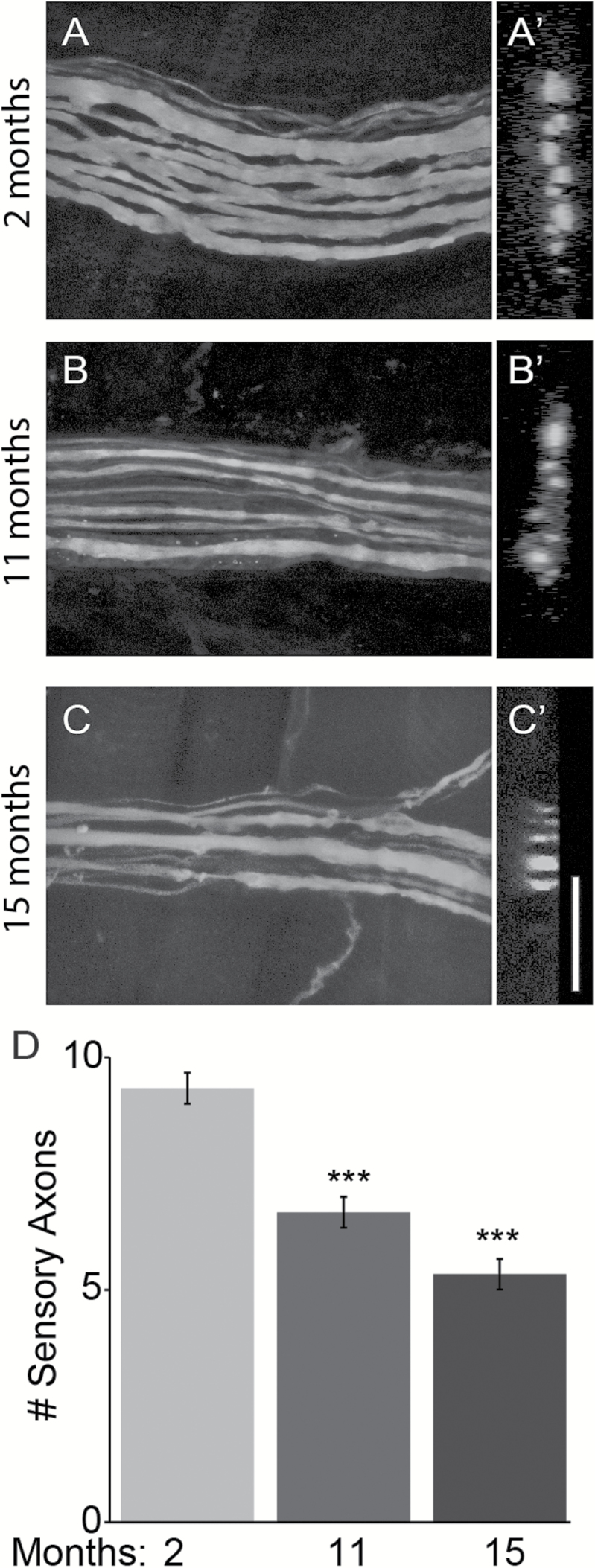
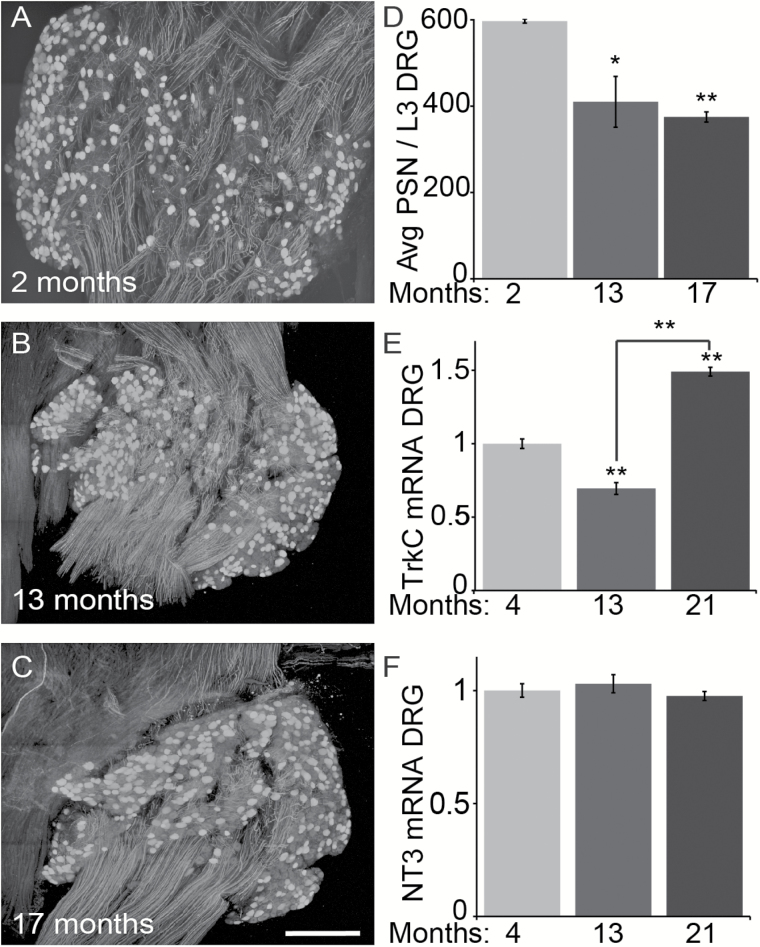
The impact of aging on proprioceptive sensory neurons and intrafusal muscle fibers (IMFs) remains largely unexplored despite the central function these cells play in modulating voluntary movements. Here, we show that proprioceptive sensory neurons undergo deleterious morphological changes in middle age (11- to 13-month-old) and old (15- to 21-month-old) mice. In the extensor digitorum longus and soleus muscles of middle age and old mice, there is a significant increase in the number of Ia afferents with large swellings that fail to properly wrap around IMFs compared with young adult (2- to 4-month-old) mice. Fewer II afferents were also found in the same muscles of middle age and old mice. Although these age-related changes in peripheral nerve endings were accompanied by degeneration of proprioceptive sensory neuron cell bodies in dorsal root ganglia (DRG), the morphology and number of IMFs remained unchanged. Our analysis also revealed normal levels of neurotrophin 3 (NT3) but dysregulated expression of the tyrosine kinase receptor C (TrkC) in aged muscles and DRGs, respectively. These results show that proprioceptive sensory neurons degenerate prior to atrophy of IMFs during aging, and in the presence of the NT3/TrkC signaling axis.
Keywords: Degeneration; Dorsal root ganglia; Ia/II afferents; Motor function; Muscle spindle.
© The Author 2016. Published by Oxford University Press on behalf of The Gerontological Society of America. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Fried LP, Ferrucci L, Darer J, Williamson JD, Anderson G. Untangling the concepts of disability, frailty, and comorbidity: implications for improved targeting and care. J Gerontol A Biol Sci Med Sci. 2004;59:255–263. doi:10.1093/gerona/59.3.M255 - PubMed
-
- Martin FC. Falls risk factors: assessment and management to prevent falls and fractures. Can J Aging. 2011;30:33–44. doi:10.1017/S0714980810000747 - PubMed
-
- Ambrose AF, Paul G, Hausdorff JM. Risk factors for falls among older adults: a review of the literature. Maturitas. 2013;75:51–61. doi:10.1016/j.maturitas.2013.02.009 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
