

Chemical Proteomic Profiling of Human Methyltransferases
- PMID: 27689866
- PMCID: PMC5072128
- DOI: 10.1021/jacs.6b07830
Chemical Proteomic Profiling of Human Methyltransferases
Abstract
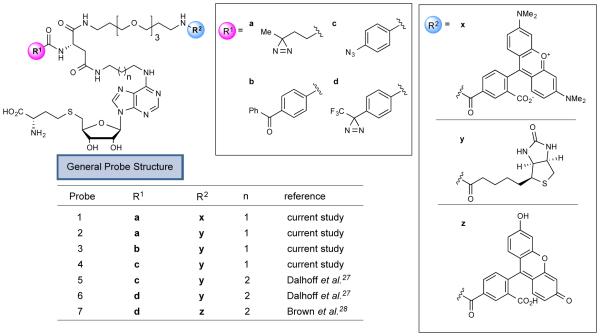
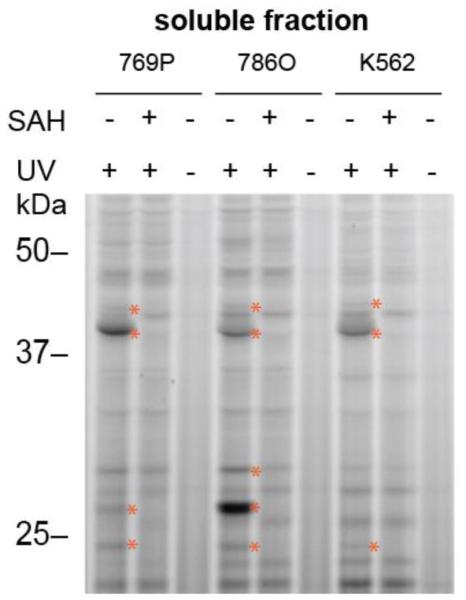
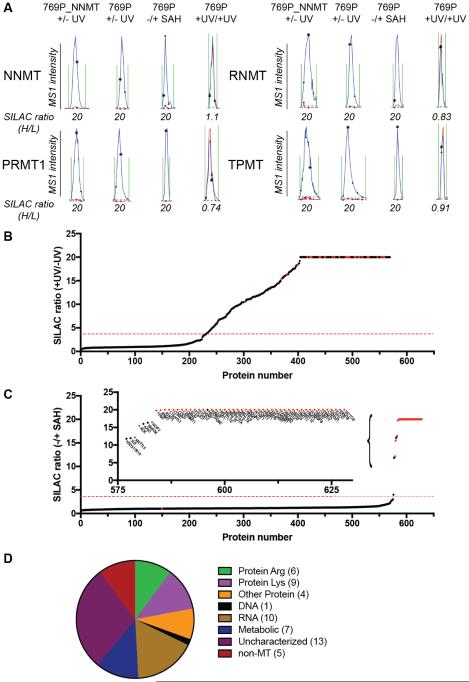
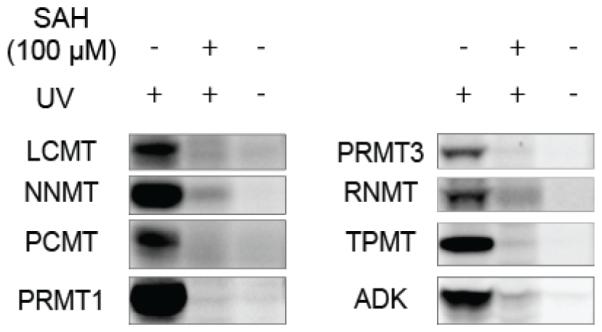
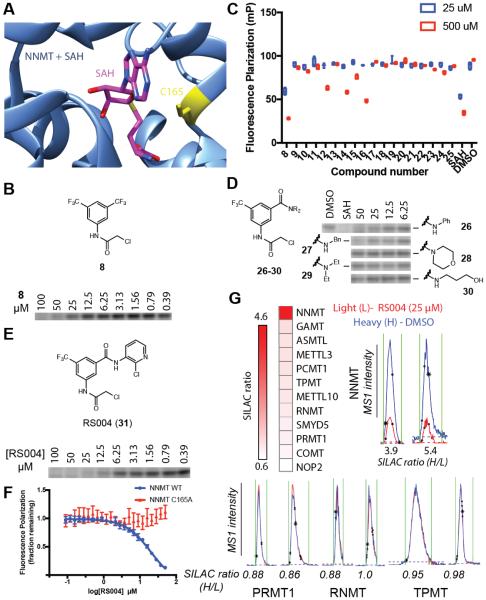
Methylation is a fundamental mechanism used in Nature to modify the structure and function of biomolecules, including proteins, DNA, RNA, and metabolites. Methyl groups are predominantly installed into biomolecules by a large and diverse class of S-adenosyl methionine (SAM)-dependent methyltransferases (MTs), of which there are ∼200 known or putative members in the human proteome. Deregulated MT activity contributes to numerous diseases, including cancer, and several MT inhibitors are in clinical development. Nonetheless, a large fraction of the human MT family remains poorly characterized, underscoring the need for new technologies to characterize MTs and their inhibitors in native biological systems. Here, we describe a suite of S-adenosyl homocysteine (SAH) photoreactive probes and their application in chemical proteomic experiments to profile and enrich a large number of MTs (>50) from human cancer cell lysates with remarkable specificity over other classes of proteins. We further demonstrate that the SAH probes can enrich MT-associated proteins and be used to screen for and assess the selectivity of MT inhibitors, leading to the discovery of a covalent inhibitor of nicotinamide N-methyltransferase (NNMT), an enzyme implicated in cancer and metabolic disorders. The chemical proteomics probes and methods for their utilization reported herein should prove of value for the functional characterization of MTs, MT complexes, and MT inhibitors in mammalian biology and disease.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
