

Protein cysteine oxidation in redox signaling: Caveats on sulfenic acid detection and quantification
- PMID: 27693037
- PMCID: PMC5318241
- DOI: 10.1016/j.abb.2016.09.013
Protein cysteine oxidation in redox signaling: Caveats on sulfenic acid detection and quantification
Abstract
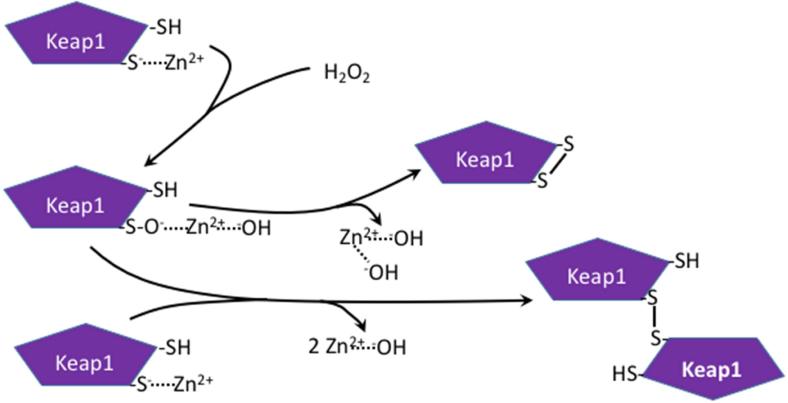
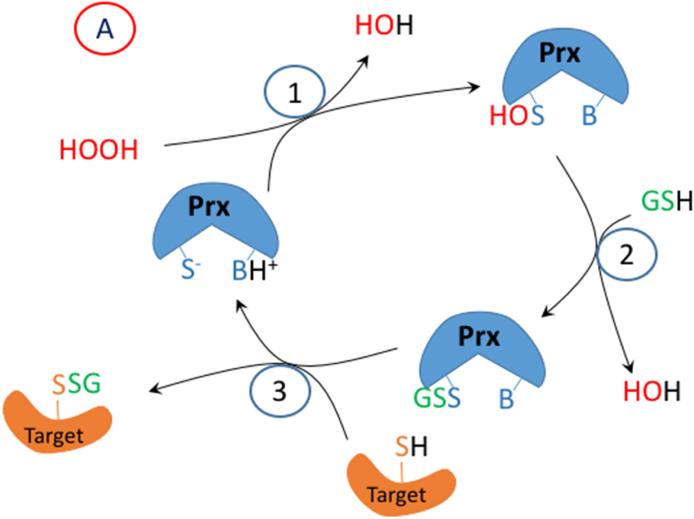
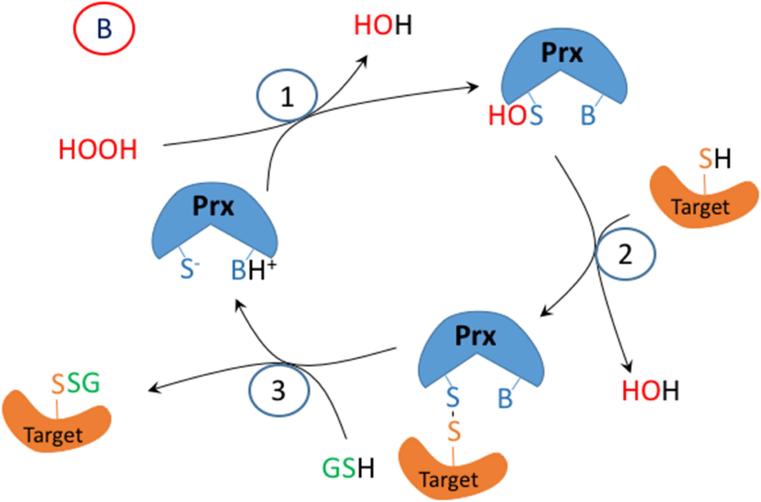
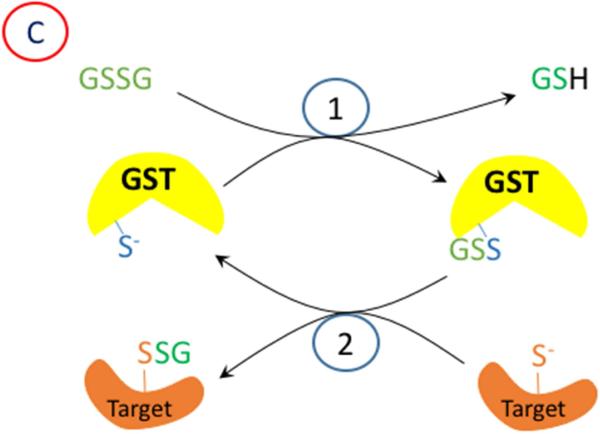
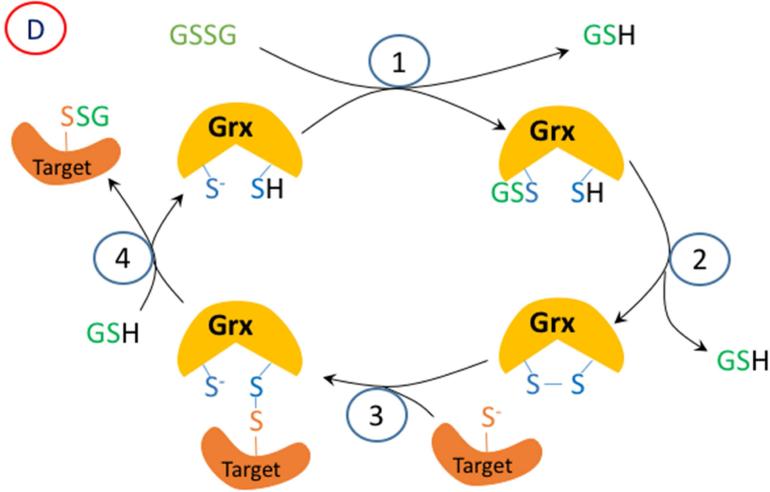
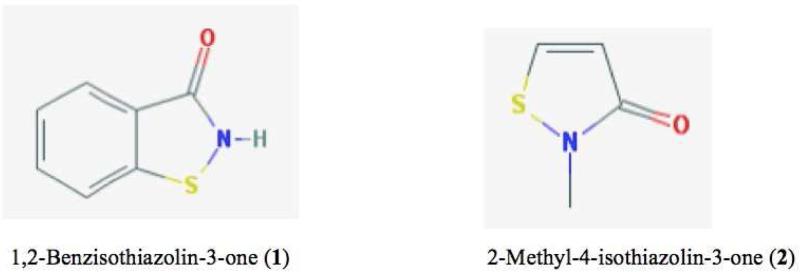
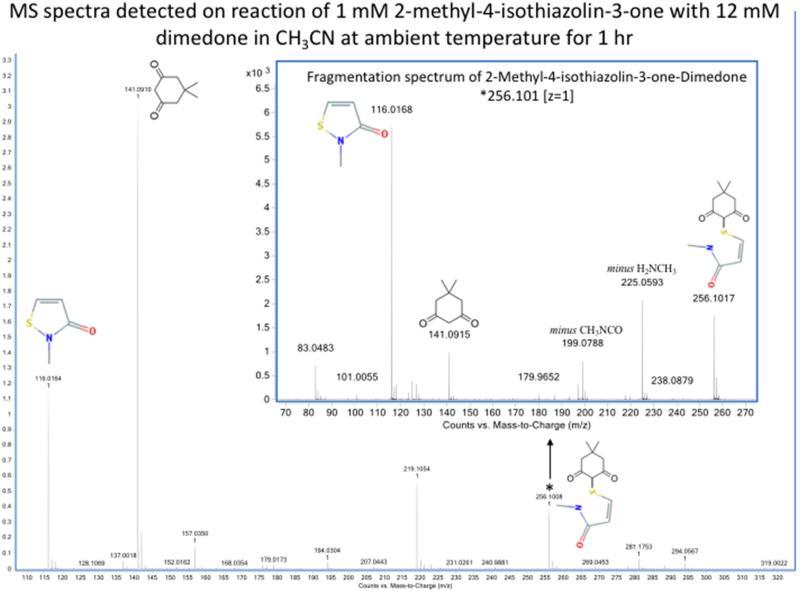
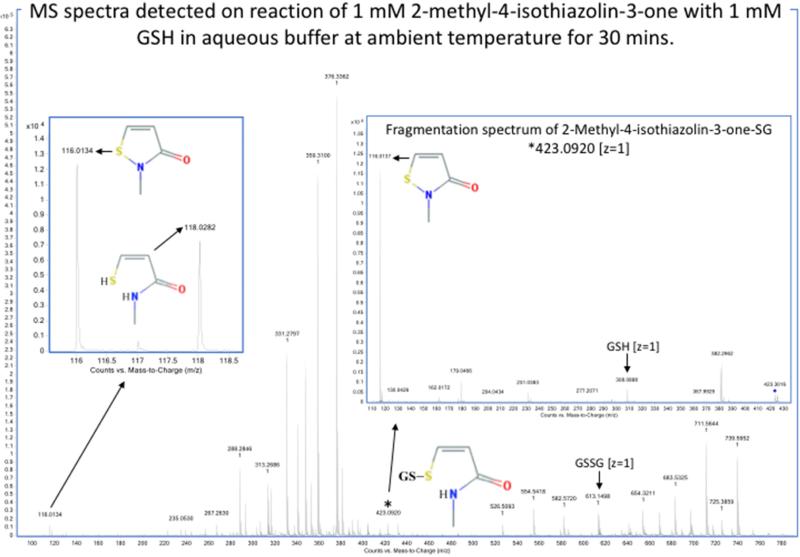
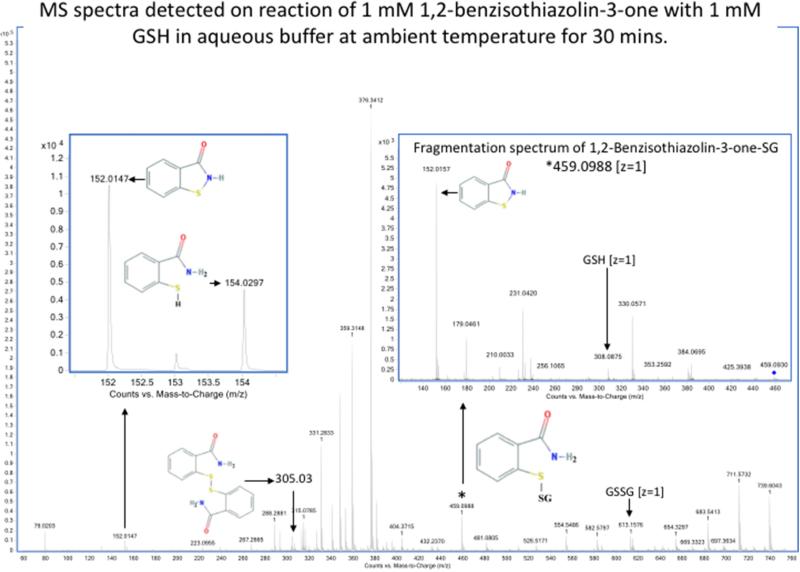
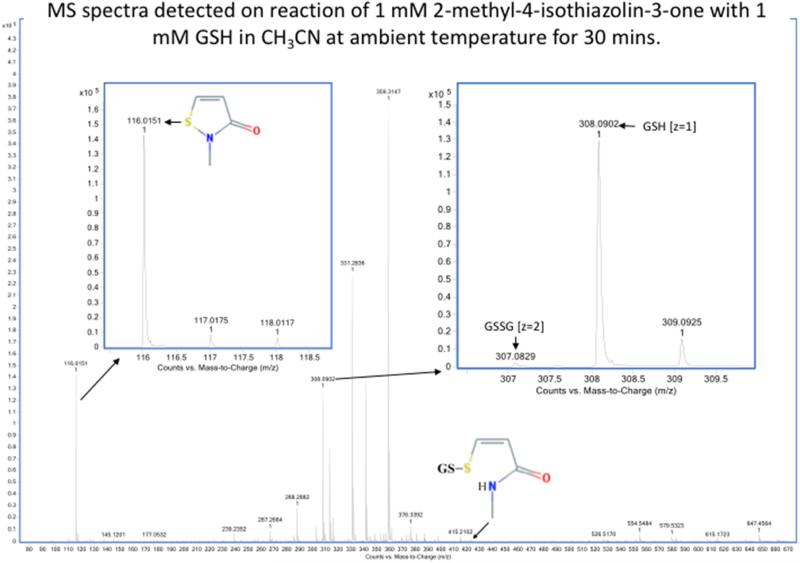
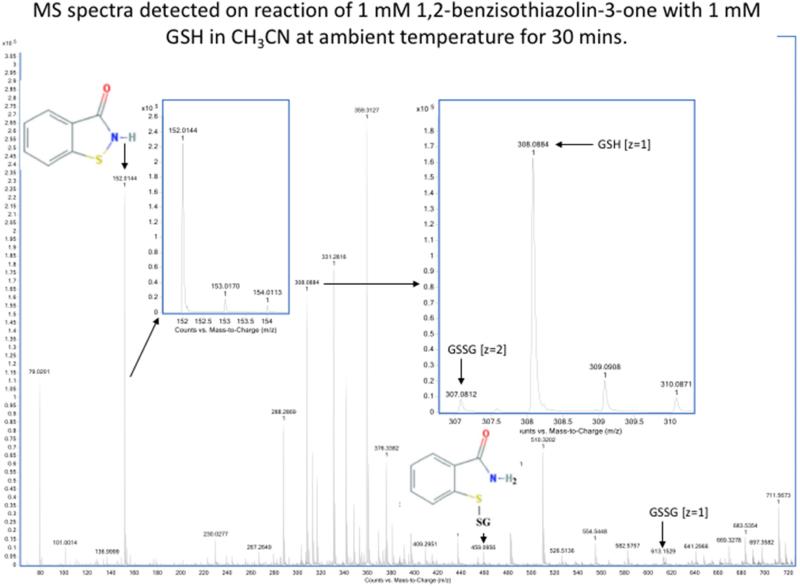
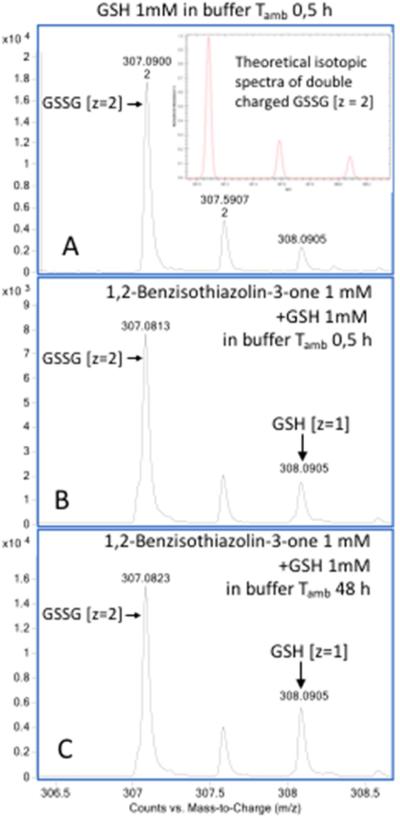
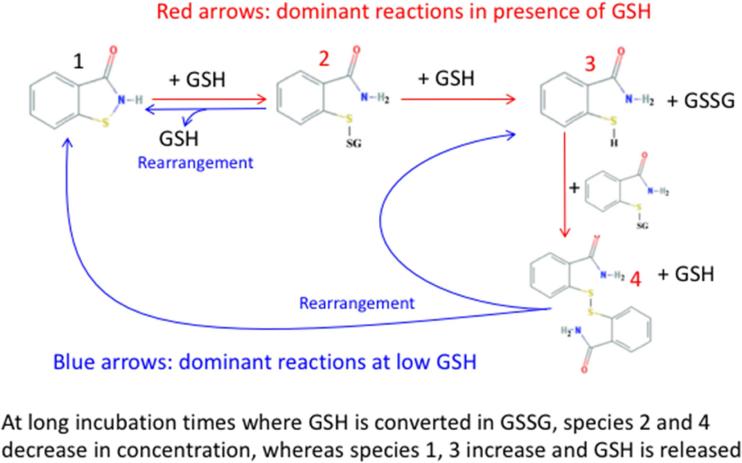
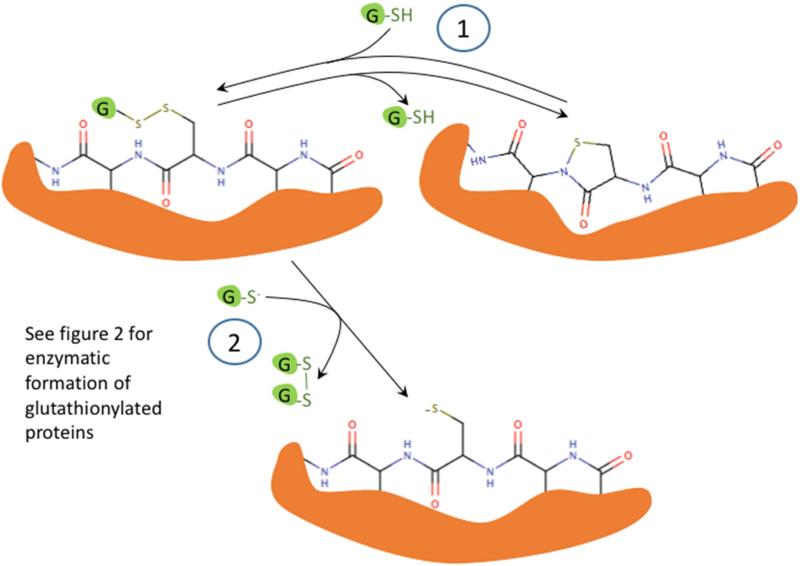
Oxidation of critical signaling protein cysteines regulated by H2O2 has been considered to involve sulfenic acid (RSOH) formation. RSOH may subsequently form either a sulfenyl amide (RSNHR') with a neighboring amide, or a mixed disulfide (RSSR') with another protein cysteine or glutathione. Previous studies have claimed that RSOH can be detected as an adduct (e.g., with 5,5-dimethylcyclohexane-1,3-dione; dimedone). Here, kinetic data are discussed which indicate that few proteins can form RSOH under physiological signaling conditions. We also present experimental evidence that indicates that (1) dimedone reacts rapidly with sulfenyl amides, and more rapidly than with sulfenic acids, and (2) that disulfides can react reversibly with amides to form sulfenyl amides. As some proteins are more stable as the sulfenyl amide than as a glutathionylated species, the former may account for some of the species previously identified as the "sulfenome" - the cellular complement of reversibly-oxidized thiol proteins generated via sulfenic acids.
Keywords: Glutathione; Hydrogen peroxide; Redox signaling; Sulfenyl amide; Thiolate.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Trujillo M, Alvarez B, Radi R. One- and two-electron oxidation of thiols: mechanisms, kinetics and biological fates. Free Radic. Res. 2016;50:150–171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
