

Hunger-Driven Motivational State Competition
- PMID: 27693254
- PMCID: PMC5082717
- DOI: 10.1016/j.neuron.2016.08.032
Hunger-Driven Motivational State Competition
Abstract
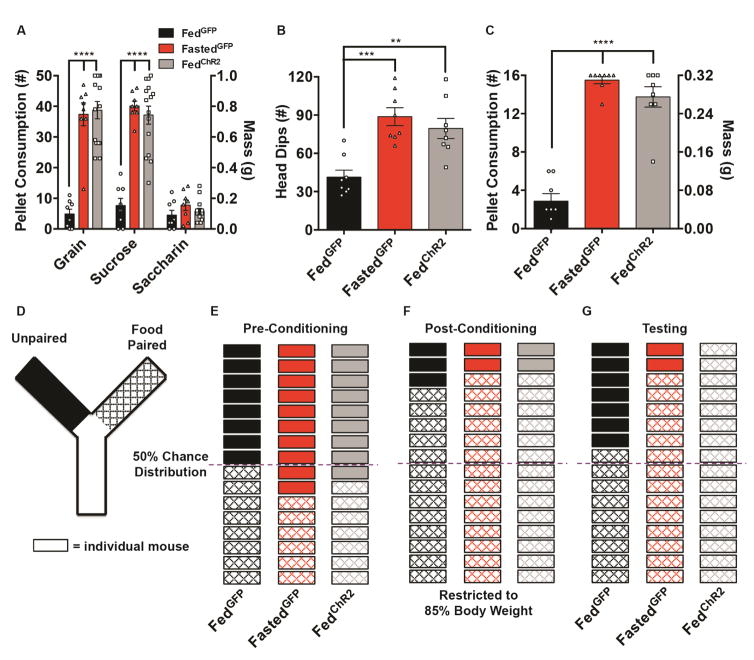
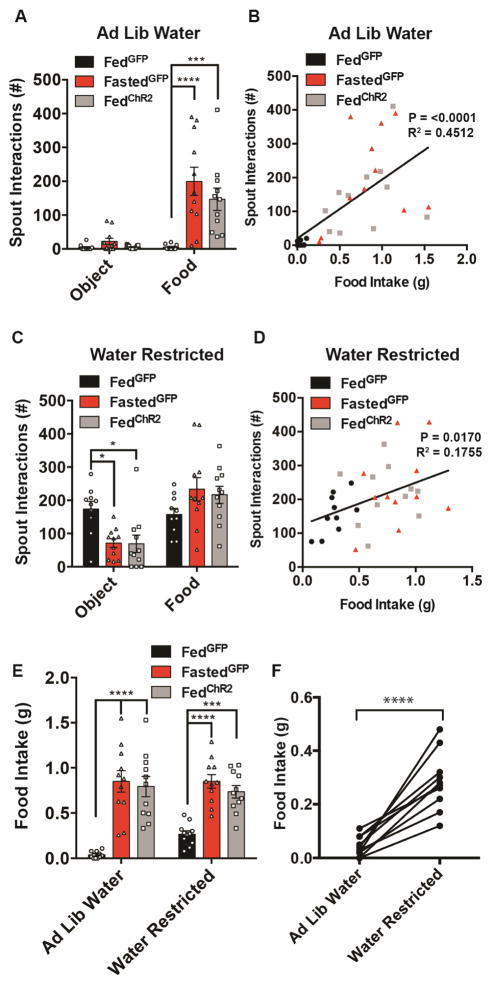
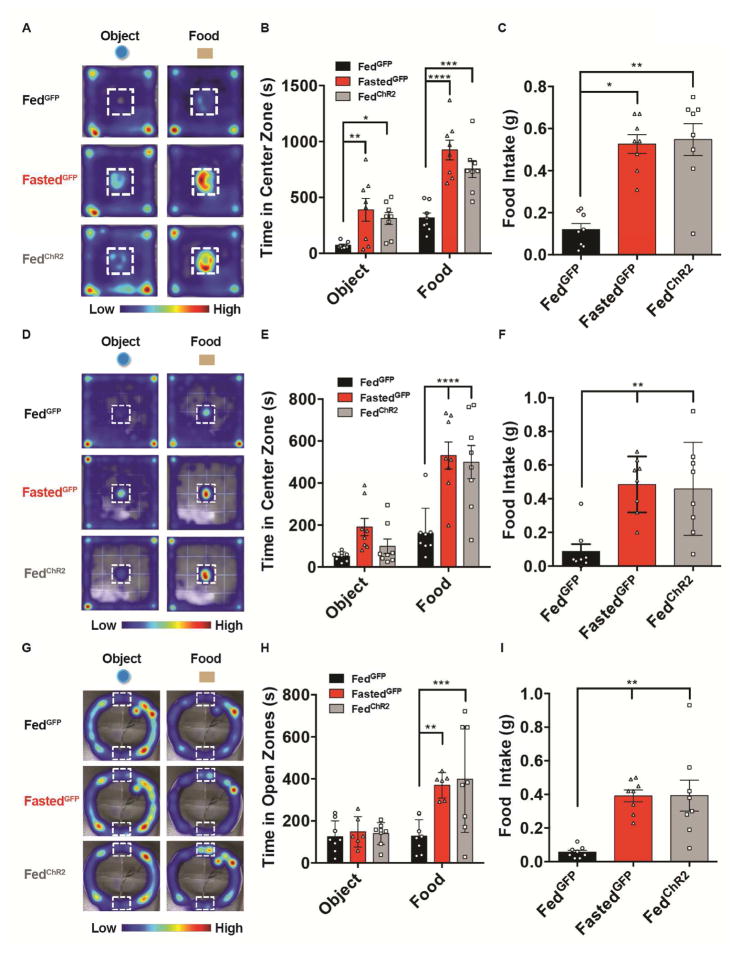
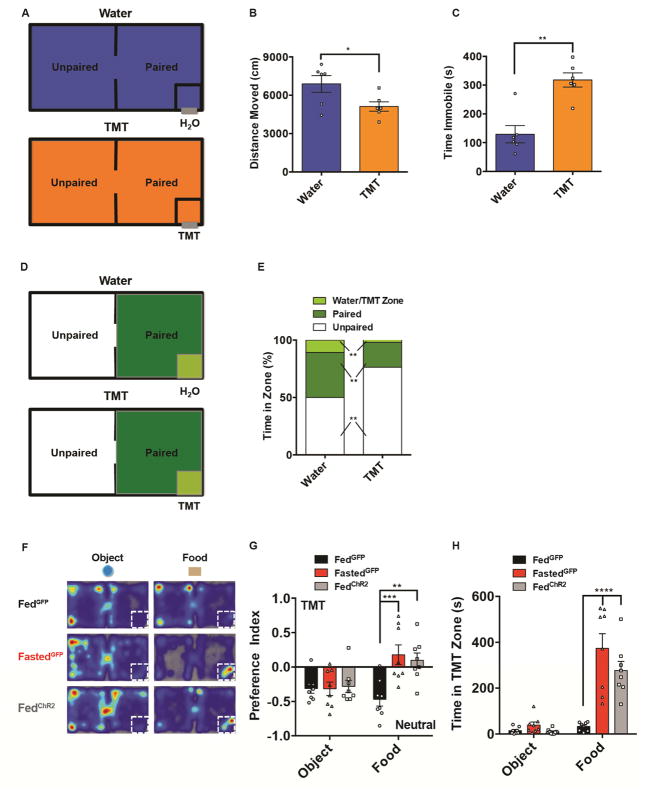
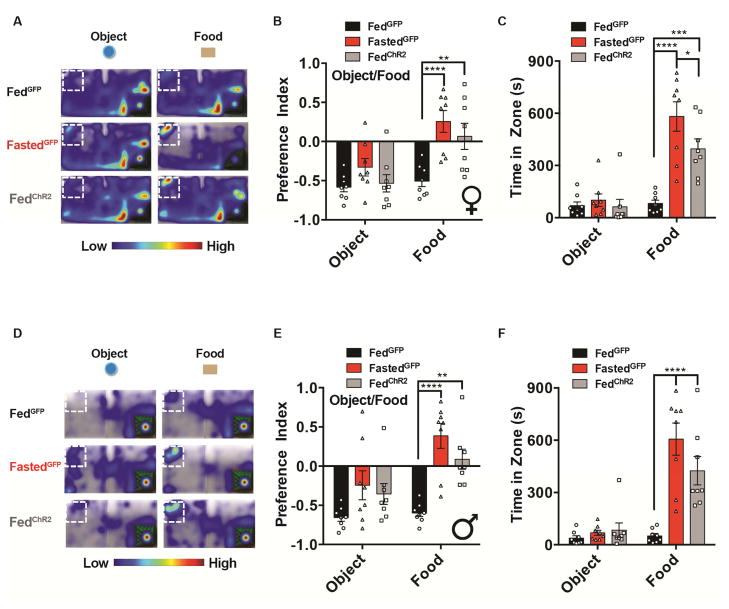
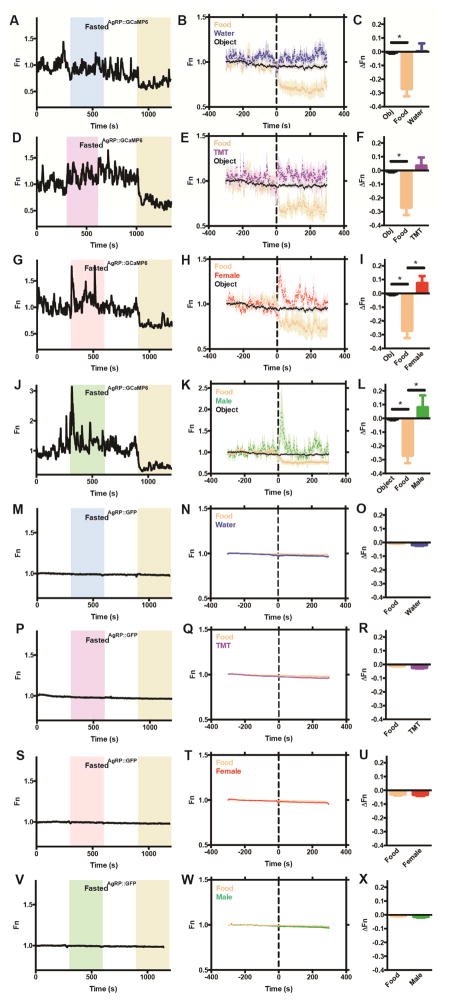
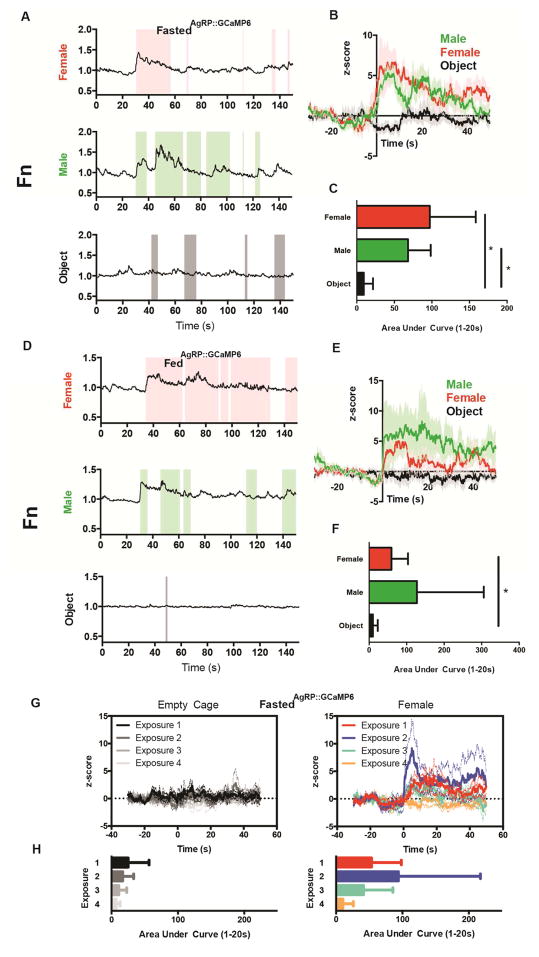
Behavioral choice is ubiquitous in the animal kingdom and is central to goal-oriented behavior. Hypothalamic Agouti-related peptide (AgRP) neurons are critical regulators of appetite. Hungry animals, bombarded by multiple sensory stimuli, are known to modify their behavior during times of caloric need, rapidly adapting to a consistently changing environment. Utilizing ARCAgRP neurons as an entry point, we analyzed the hierarchical position of hunger related to rival drive states. Employing a battery of behavioral assays, we found that hunger significantly increases its capacity to suppress competing motivational systems, such as thirst, anxiety-related behavior, innate fear, and social interactions, often only when food is accessible. Furthermore, real-time monitoring of ARCAgRP activity revealed time-locked responses to conspecific investigation in addition to food presentation, further establishing that, even at the level of ARCAgRP neurons, choices are remarkably flexible computations, integrating internal state, external factors, and anticipated yield. VIDEO ABSTRACT.
Published by Elsevier Inc.
Figures
References
-
- Berridge KC, Robinson TE. What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res Brain Res Rev. 1998;28:309–369. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
