

Zika Virus Infection Induces Cranial Neural Crest Cells to Produce Cytokines at Levels Detrimental for Neurogenesis
- PMID: 27693308
- PMCID: PMC5113290
- DOI: 10.1016/j.chom.2016.09.006
Zika Virus Infection Induces Cranial Neural Crest Cells to Produce Cytokines at Levels Detrimental for Neurogenesis
Abstract
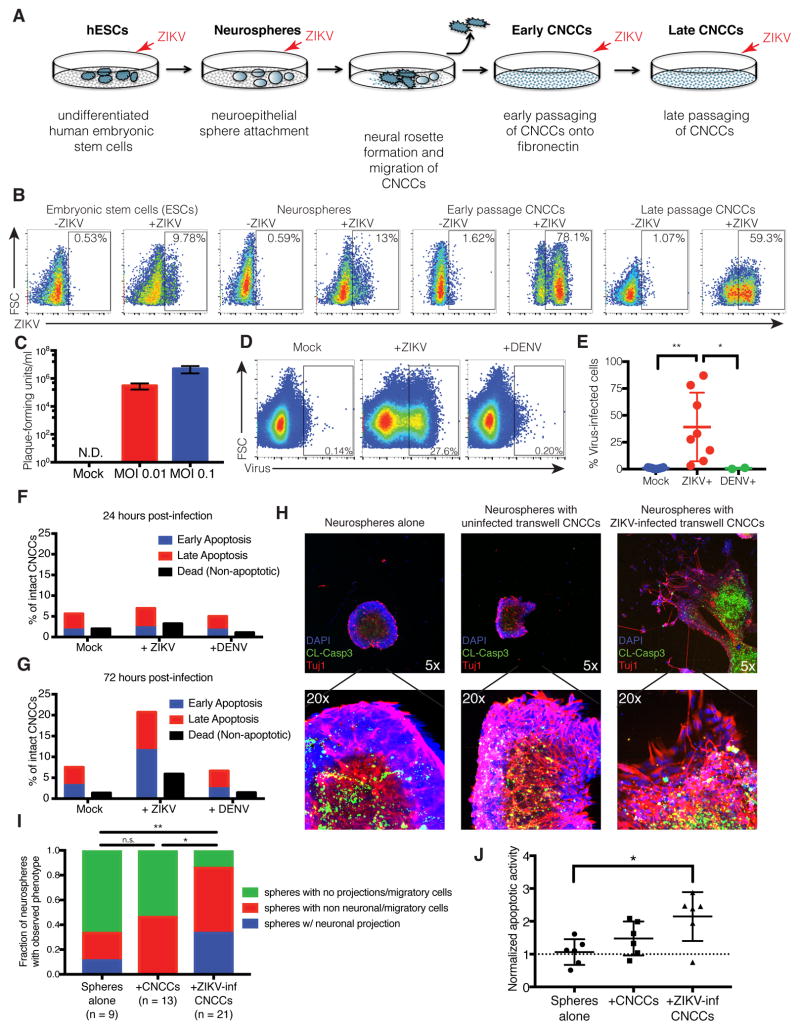
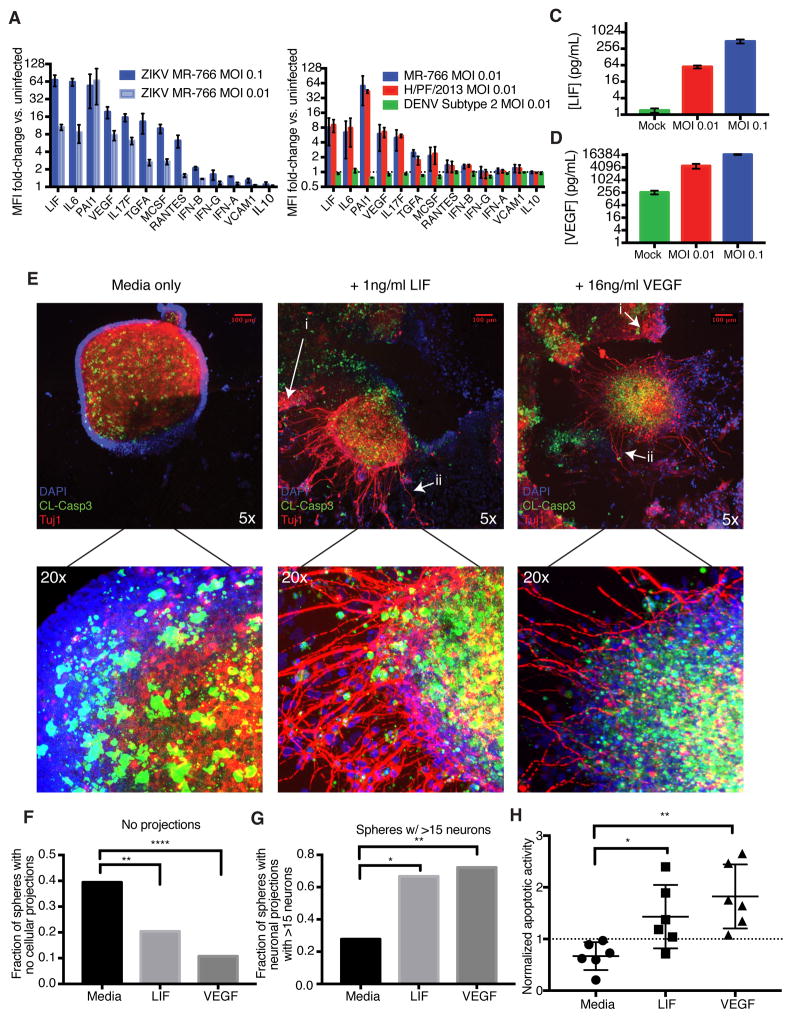
Zika virus (ZIKV) infection during pregnancy is linked to microcephaly, which is attributed to infection of developing brain structures. ZIKV infects neural progenitor cells in vitro, though its effects on other developmentally relevant stem cell populations, including cranial neural crest cells (CNCCs), have not been assessed. CNCCs give rise to most cranial bones and exert paracrine effects on the developing brain. Here, we report that CNCCs are productively infected by ZIKV, but not by the related dengue virus. ZIKV-infected CNCCs undergo limited apoptosis but secrete cytokines that promote death and drive aberrant differentiation of neural progenitor cultures. Addition of two such cytokines, LIF or VEGF, at levels comparable to those secreted by ZIKV-infected CNCCs is sufficient to recapitulate premature neuronal differentiation and apoptotic death of neural progenitors. Thus, our results suggest that CNCC infection by ZIKV may contribute to associated embryopathies through signaling crosstalk between developing face and brain structures.
Keywords: Zika virus; cytokines; growth factors; microcephaly; neural development; stem cells.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Example of non-neuronal migratory cellular outgrowth
Example of neuronal projections
Examples of non-neuronal migratory cellular outgrowth
Examples of neuronal outgrowth
References
-
- Brasil P, Pereira JP, Raja Gabaglia C, Damasceno L, Wakimoto M, Ribeiro Nogueira RM, Carvalho de Sequeira P, Machado Siqueira A, Abreu de Carvalho LM, Cotrim da Cunha D, Calvet GA, Neves ES, Moreira ME, Rodrigues Baião AE, Nassar de Carvalho PR, Janzen C, Valderramos SG, Cherry JD, Bispo de Filippis AM, Nielsen-Saines K. Zika Virus Infection in Pregnant Women in Rio de Janeiro - Preliminary Report. new england Journal of Medicine. 2016 doi: 10.1056/NEJMoa1602412. - DOI - PMC - PubMed
-
- de Fatima Vasco Aragao M, van der Linden V, Brainer-Lima AM, Coeli RR, Rocha MA, Sobral da Silva P, Durce Costa Gomes de Carvalho M, van der Linden A, Cesario de Holanda A, Valenca MM. Clinical features and neuroimaging (CT and MRI) findings in presumed Zika virus related congenital infection and microcephaly: retrospective case series study. BMJ. 2016;353:i1901. doi: 10.1136/bmj.i1901. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
