

Loss of the HVEM Tumor Suppressor in Lymphoma and Restoration by Modified CAR-T Cells
- PMID: 27693350
- PMCID: PMC5221752
- DOI: 10.1016/j.cell.2016.08.032
Loss of the HVEM Tumor Suppressor in Lymphoma and Restoration by Modified CAR-T Cells
Abstract
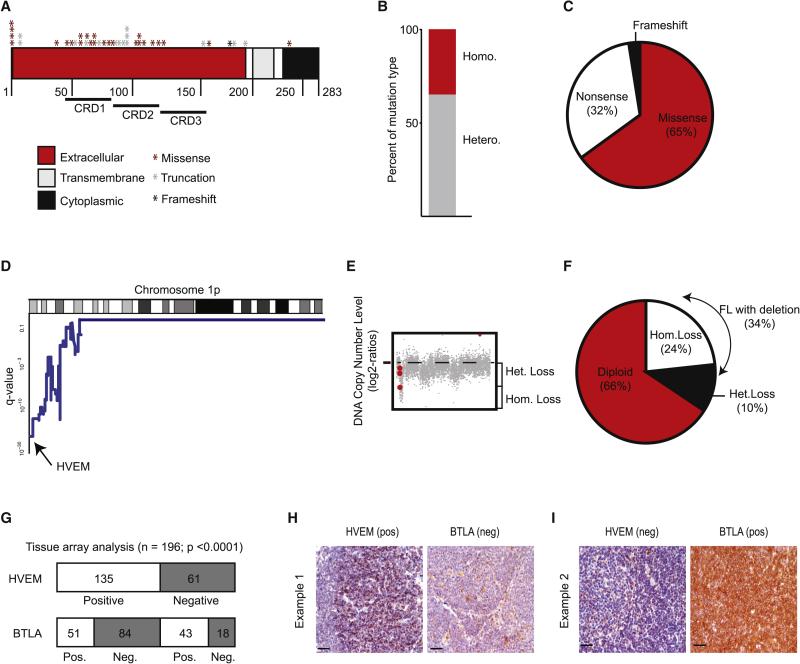
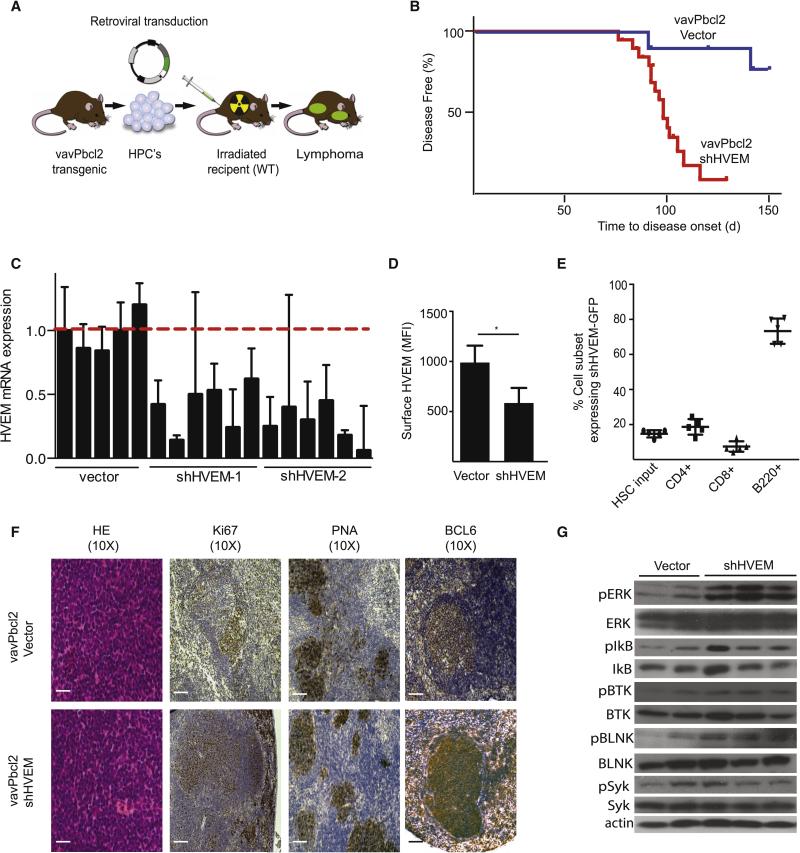
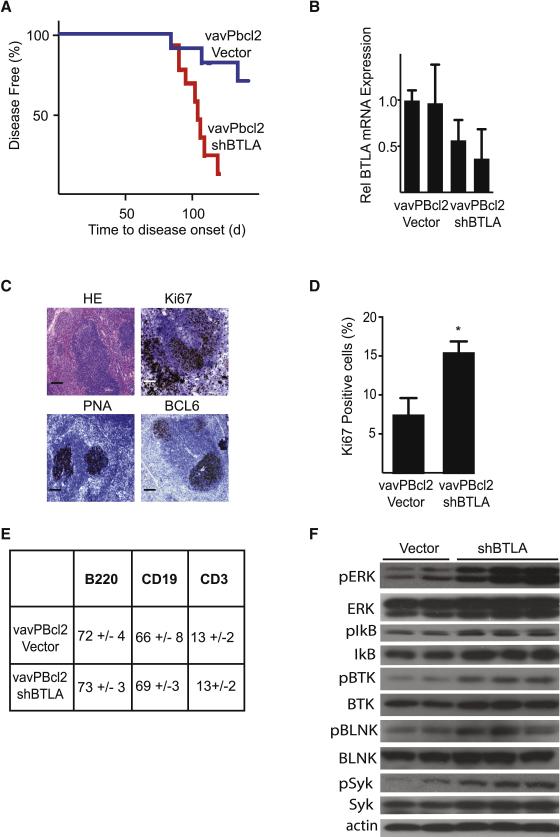
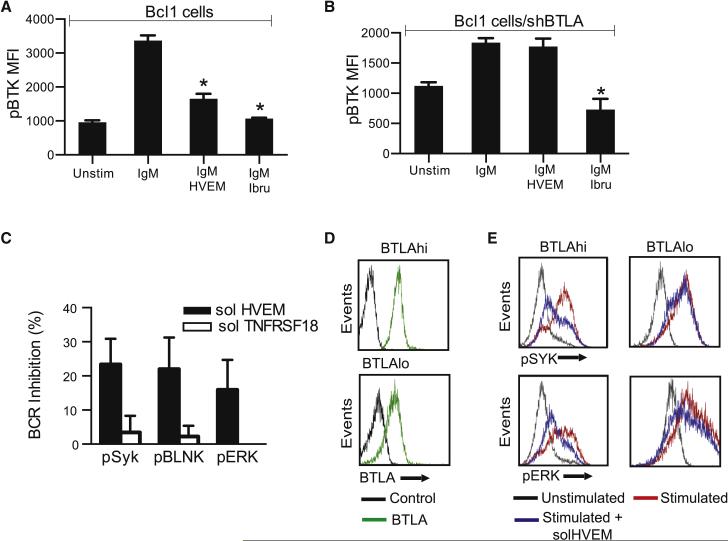
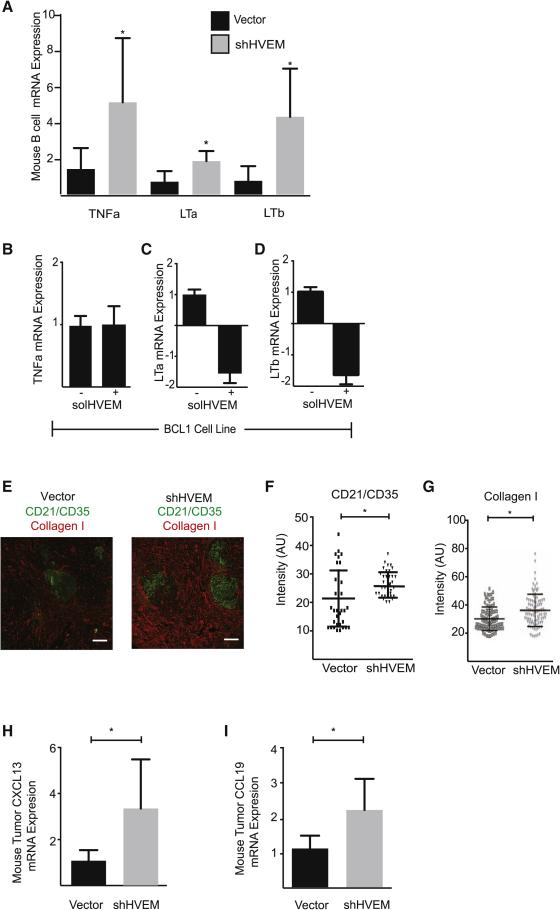
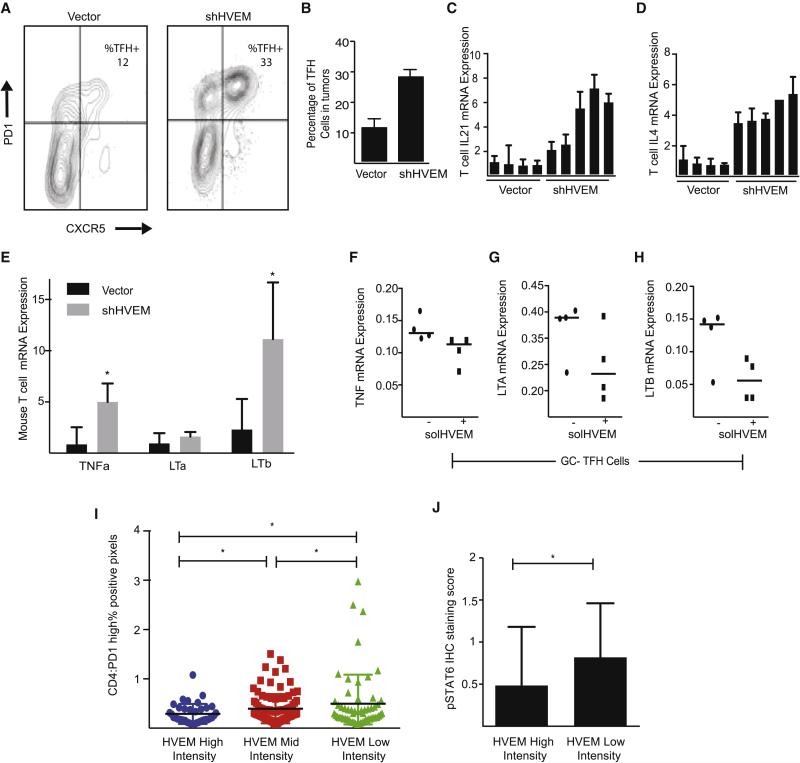
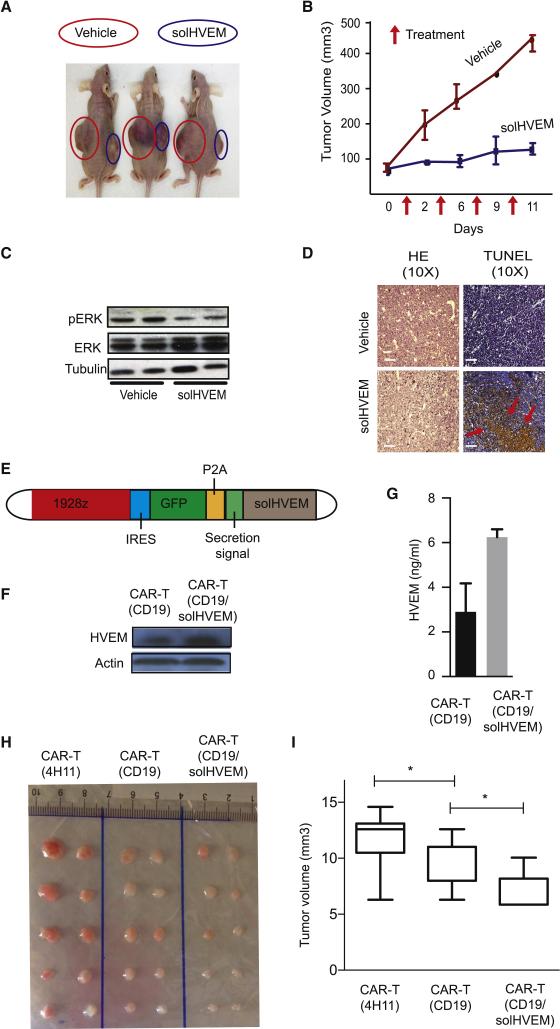
The HVEM (TNFRSF14) receptor gene is among the most frequently mutated genes in germinal center lymphomas. We report that loss of HVEM leads to cell-autonomous activation of B cell proliferation and drives the development of GC lymphomas in vivo. HVEM-deficient lymphoma B cells also induce a tumor-supportive microenvironment marked by exacerbated lymphoid stroma activation and increased recruitment of T follicular helper (TFH) cells. These changes result from the disruption of inhibitory cell-cell interactions between the HVEM and BTLA (B and T lymphocyte attenuator) receptors. Accordingly, administration of the HVEM ectodomain protein (solHVEM(P37-V202)) binds BTLA and restores tumor suppression. To deliver solHVEM to lymphomas in vivo, we engineered CD19-targeted chimeric antigen receptor (CAR) T cells that produce solHVEM locally and continuously. These modified CAR-T cells show enhanced therapeutic activity against xenografted lymphomas. Hence, the HVEM-BTLA axis opposes lymphoma development, and our study illustrates the use of CAR-T cells as "micro-pharmacies" able to deliver an anti-cancer protein.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Customizing Functionality and Payload Delivery for Receptor-Engineered T Cells.Cell. 2016 Oct 6;167(2):304-306. doi: 10.1016/j.cell.2016.09.033. Cell. 2016. PMID: 27716501 Free PMC article.
References
-
- Aguzzi A, Kranich J, Krautler NJ. Follicular dendritic cells: origin, phenotype, and function in health and disease. Trends Immunol. 2014;35:105–113. - PubMed
-
- Ame-Thomas P, Maby-El Hajjami H, Monvoisin C, Jean R, Monnier D, Caulet-Maugendre S, Guillaudeux T, Lamy T, Fest T, Tarte K. Human mesenchymal stem cells isolated from bone marrow and lymphoid organs support tumor B-cell growth: role of stromal cells in follicular lymphoma pathogenesis. Blood. 2007;109:693–702. - PubMed
-
- Ame-Thomas P, Tarte K. The yin and the yang of follicular lymphoma cell niches: role of microenvironment heterogeneity and plasticity. Semin Cancer Biol. 2014;24:23–32. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
