

Replication-Dependent Unhooking of DNA Interstrand Cross-Links by the NEIL3 Glycosylase
- PMID: 27693351
- PMCID: PMC5237264
- DOI: 10.1016/j.cell.2016.09.008
Replication-Dependent Unhooking of DNA Interstrand Cross-Links by the NEIL3 Glycosylase
Abstract
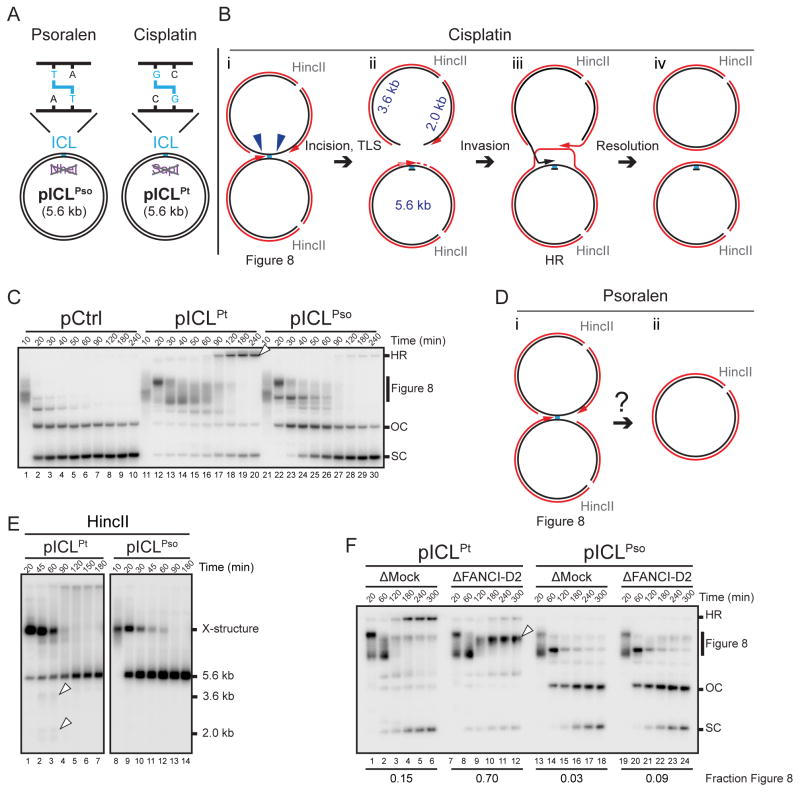
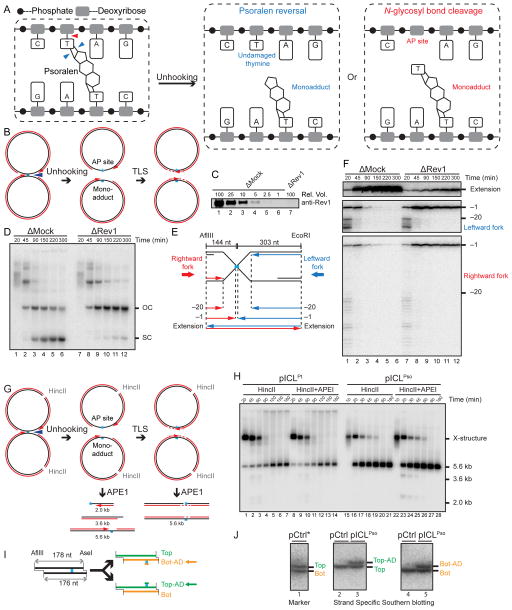
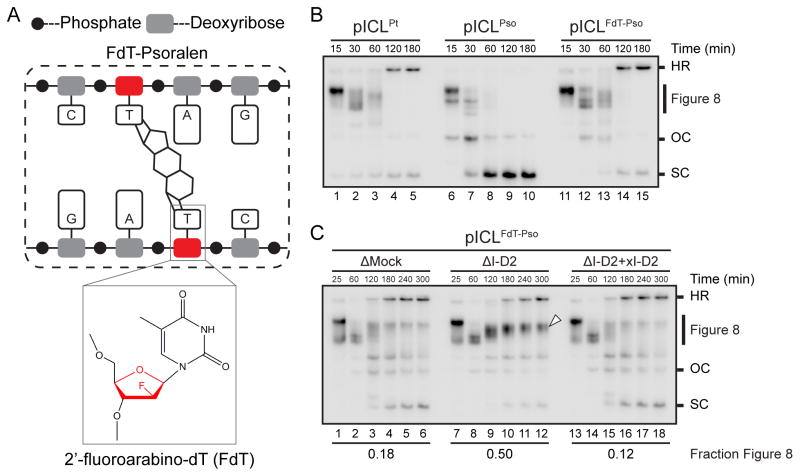
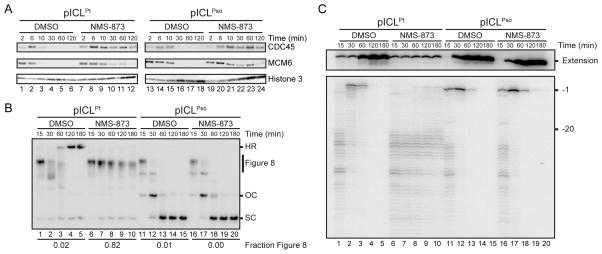
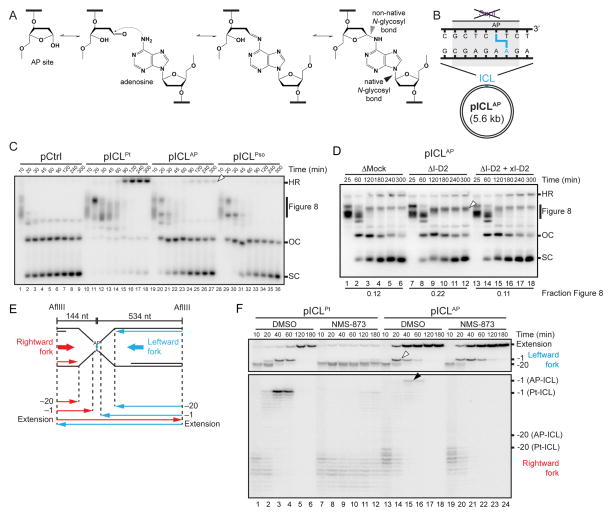
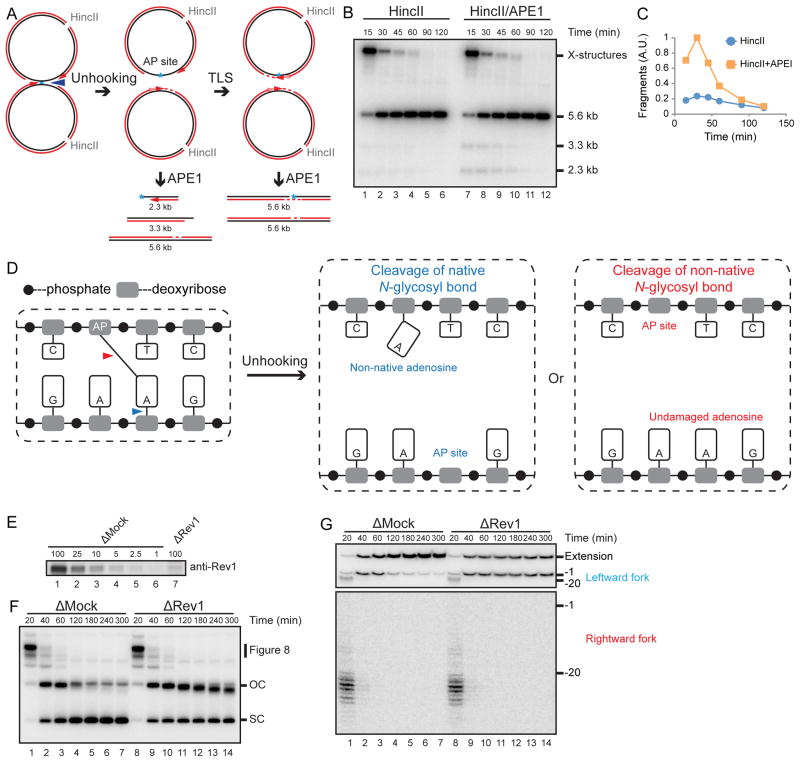
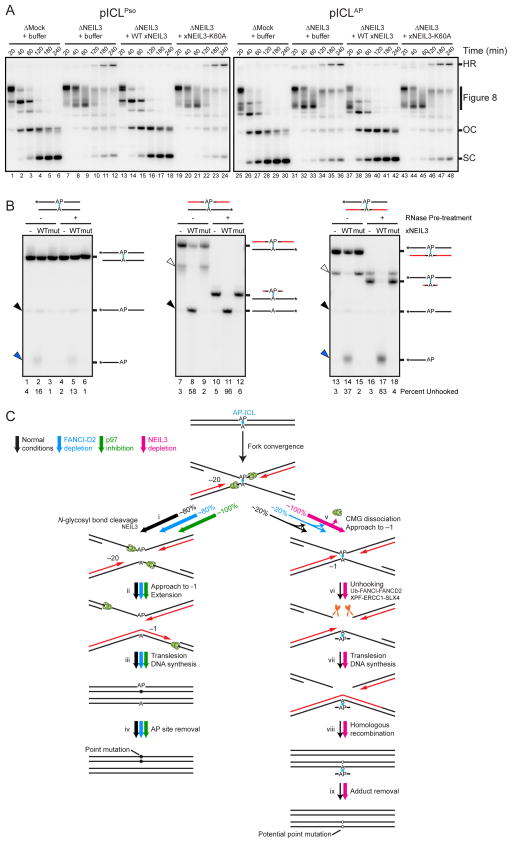
During eukaryotic DNA interstrand cross-link (ICL) repair, cross-links are resolved ("unhooked") by nucleolytic incisions surrounding the lesion. In vertebrates, ICL repair is triggered when replication forks collide with the lesion, leading to FANCI-FANCD2-dependent unhooking and formation of a double-strand break (DSB) intermediate. Using Xenopus egg extracts, we describe here a replication-coupled ICL repair pathway that does not require incisions or FANCI-FANCD2. Instead, the ICL is unhooked when one of the two N-glycosyl bonds forming the cross-link is cleaved by the DNA glycosylase NEIL3. Cleavage by NEIL3 is the primary unhooking mechanism for psoralen and abasic site ICLs. When N-glycosyl bond cleavage is prevented, unhooking occurs via FANCI-FANCD2-dependent incisions. In summary, we identify an incision-independent unhooking mechanism that avoids DSB formation and represents the preferred pathway of ICL repair in a vertebrate cell-free system.
Keywords: DNA interstrand cross-link; FANCD2; FANCI; Fanconi anemia; ICL; NEIL3; abasic site; psoralen.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Alt A, Lammens K, Chiocchini C, Lammens A, Pieck JC, Kuch D, Hopfner KP, Carell T. Bypass of DNA lesions generated during anticancer treatment with cisplatin by DNA polymerase eta. Science. 2007;318:967–970. - PubMed
-
- Cimino GD, Shi YB, Hearst JE. Wavelength dependence for the photoreversal of a psoralen-DNA cross-link. Biochemistry. 1986;25:3013–3020. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
