

Energetics of acclimation to NaCl by submerged, anoxic rice seedlings
- PMID: 27694332
- PMCID: PMC5218384
- DOI: 10.1093/aob/mcw189
Energetics of acclimation to NaCl by submerged, anoxic rice seedlings
Abstract
Background and aims: Our aim was to elucidate how plant tissues under a severe energy crisis cope with imposition of high NaCl, which greatly increases ion fluxes and hence energy demands. The energy requirements for ion regulation during combined salinity and anoxia were assessed to gain insights into ion transport processes in the anoxia-tolerant coleoptile of rice.
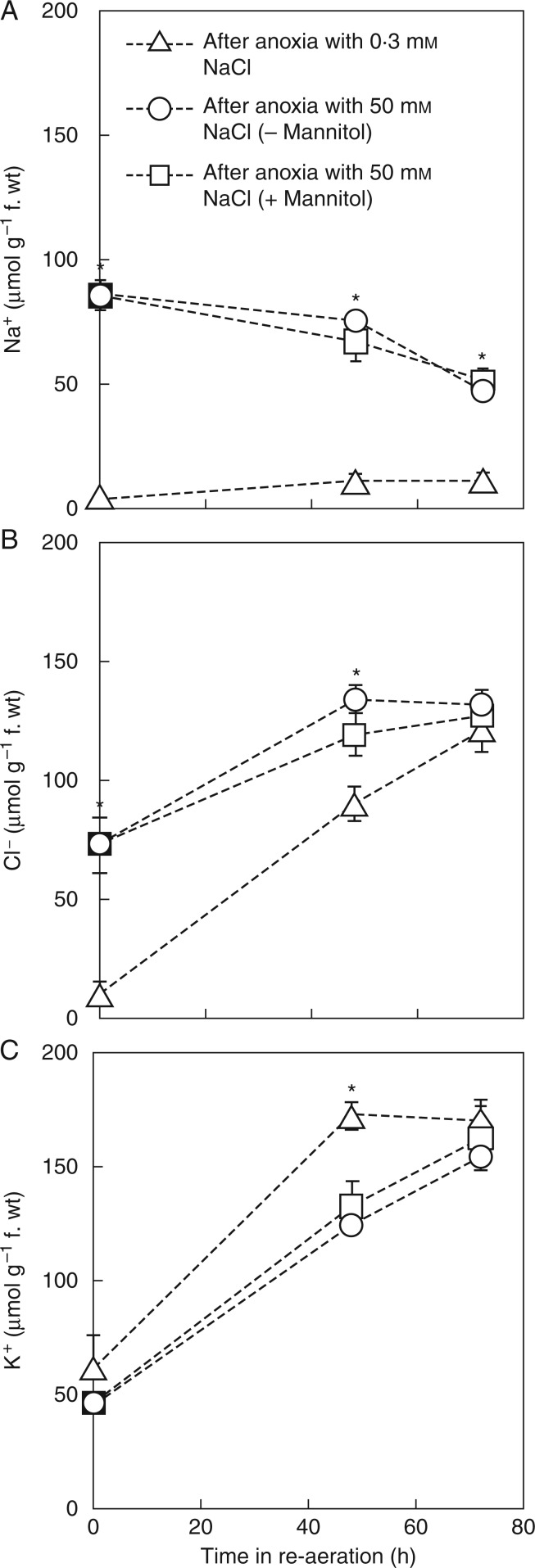
Methods: We studied the combined effects of anoxia plus 50 or 100 mm NaCl on tissue ions and growth of submerged rice (Oryza sativa) seedlings. Excised coleoptiles allowed measurements in aerated or anoxic conditions of ion net fluxes and O2 consumption or ethanol formation and by inference energy production.
Key results: Over 80 h of anoxia, coleoptiles of submerged intact seedlings grew at 100 mm NaCl, but excised coleoptiles, with 50 mm exogenous glucose, survived only at 50 mm NaCl, possibly due to lower energy production with glucose than for intact coleoptiles with sucrose as substrate. Rates of net uptake of Na+ and Cl- by coleoptiles in anoxia were about half those in aerated solution. Ethanol formation in anoxia and O2 uptake in aerobic solution were each increased by 13-15 % at 50 mm NaCl, i.e. ATP formation was stimulated. For acclimation to 50 mm NaCl, the anoxic tissues used only 25 % of the energy that was expended by aerobic tissues. Following return of coleoptiles to aerated non-saline solution, rates of net K+ uptake recovered to those in continuously aerated solution, demonstrating there was little injury during anoxia with 50 mm NaCl.
Conclusion: Rice seedlings survive anoxia, without the coleoptile incurring significant injury, even with the additional energy demands imposed by NaCl (100 mm when intact, 50 mm when excised). Energy savings were achieved in saline anoxia by less coleoptile growth, reduced ion fluxes as compared to aerobic coleoptiles and apparent energy-economic ion transport systems.
Keywords: Anoxia tolerance; NaCl ×; Oryza sativa; anaerobic catabolism; anoxia interaction; anoxia plus NaCl; coleoptile; complete submergence; energetics; energy crisis; energy efficient transport; ethanolic fermentation; germination; salinity tolerance.
© The Author 2016. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures






References
-
- Atwell BJ, Water I, Greenway H. 1982. The effect of oxygen and turbulence on elongation of coleoptiles of submergence-tolerant and -intolerant rice cultivars. Journal of Experimental Botany 33: 1030–1044.
-
- Atwell BJ, Greenway H, Colmer TD. 2015. Efficient use of energy in anoxia-tolerant plants with focus on germinating rice seedlings. New Phytologist 206: 36–56. - PubMed
-
- Barrett-Lennard EG. 2003. The interaction between waterlogging and salinity in higher plants: causes, consequences and implications. Plant and Soil 253: 35–54.
-
- Baunsgaard L, Venema K, Axelsen KB, et al. 1996. Modified plasma membrane H+-ATPase with improved transport coupling identified by mutant selection in yeast. The Plant Journal 10: 451–458. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
