

Tumor Cell-Independent Estrogen Signaling Drives Disease Progression through Mobilization of Myeloid-Derived Suppressor Cells
- PMID: 27694385
- PMCID: PMC5222699
- DOI: 10.1158/2159-8290.CD-16-0502
Tumor Cell-Independent Estrogen Signaling Drives Disease Progression through Mobilization of Myeloid-Derived Suppressor Cells
Abstract
The role of estrogens in antitumor immunity remains poorly understood. Here, we show that estrogen signaling accelerates the progression of different estrogen-insensitive tumor models by contributing to deregulated myelopoiesis by both driving the mobilization of myeloid-derived suppressor cells (MDSC) and enhancing their intrinsic immunosuppressive activity in vivo Differences in tumor growth are dependent on blunted antitumor immunity and, correspondingly, disappear in immunodeficient hosts and upon MDSC depletion. Mechanistically, estrogen receptor alpha activates the STAT3 pathway in human and mouse bone marrow myeloid precursors by enhancing JAK2 and SRC activity. Therefore, estrogen signaling is a crucial mechanism underlying pathologic myelopoiesis in cancer. Our work suggests that new antiestrogen drugs that have no agonistic effects may have benefits in a wide range of cancers, independently of the expression of estrogen receptors in tumor cells, and may synergize with immunotherapies to significantly extend survival.
Significance: Ablating estrogenic activity delays malignant progression independently of the tumor cell responsiveness, owing to a decrease in the mobilization and immunosuppressive activity of MDSCs, which boosts T-cell-dependent antitumor immunity. Our results provide a mechanistic rationale to block estrogen signaling with newer antagonists to boost the effectiveness of anticancer immunotherapies. Cancer Discov; 7(1); 72-85. ©2016 AACR.See related commentary by Welte et al., p. 17This article is highlighted in the In This Issue feature, p. 1.
©2016 American Association for Cancer Research.
Conflict of interest statement
COI: The authors declare no conflict of interest
Figures
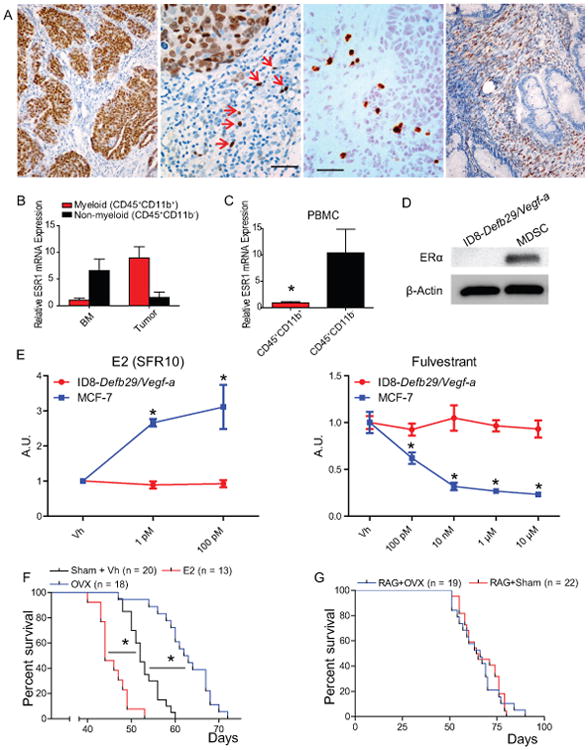


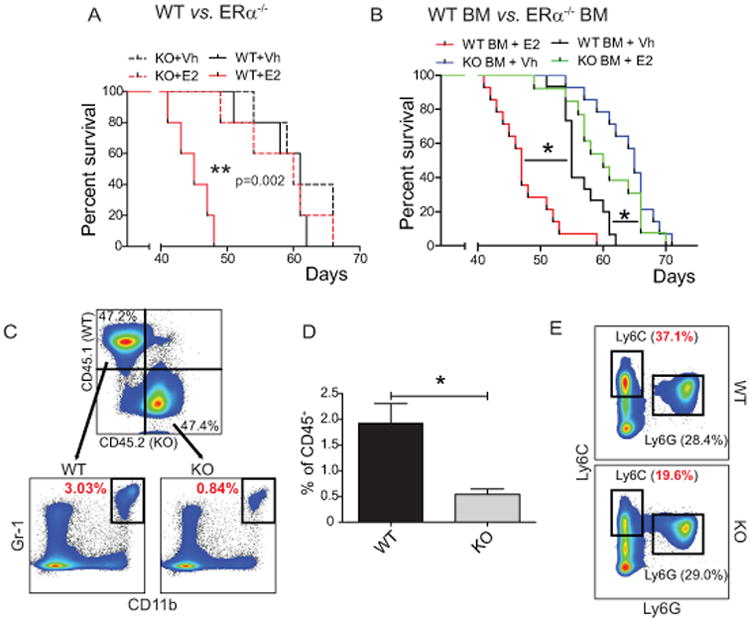



Comment in
-
Repurposing Antiestrogens for Tumor Immunotherapy.Cancer Discov. 2017 Jan;7(1):17-19. doi: 10.1158/2159-8290.CD-16-1308. Cancer Discov. 2017. PMID: 28062672 Free PMC article.
-
Tamoxifen as an immunotherapy.Sci Signal. 2017 Feb 14;10(466):eaam9611. doi: 10.1126/scisignal.aam9611. Sci Signal. 2017. PMID: 28196912
References
-
- Gallo MA, Kaufman D. Antagonistic and agonistic effects of tamoxifen: significance in human cancer. Semin Oncol. 1997;24:S1-71–S1-80. - PubMed
-
- Sini V, Cinieri S, Conte P, De Laurentiis M, Leo AD, Tondini C, et al. Endocrine therapy in post-menopausal women with metastatic breast cancer: From literature and guidelines to clinical practice. Crit Rev Oncol Hematol. 2016;100:57–68. - PubMed
-
- del Carmen MG, Fuller AF, Matulonis U, Horick NK, Goodman A, Duska LR, et al. Phase II trial of anastrozole in women with asymptomatic mullerian cancer. Gynecol Oncol. 2003;91:596–602. - PubMed
-
- Smyth JF, Gourley C, Walker G, MacKean MJ, Stevenson A, Williams AR, et al. Antiestrogen therapy is active in selected ovarian cancer cases: the use of letrozole in estrogen receptor-positive patients. Clin Cancer Res. 2007;13:3617–22. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
