

Identification of a role for TRIM29 in the control of innate immunity in the respiratory tract
- PMID: 27695001
- PMCID: PMC5558830
- DOI: 10.1038/ni.3580
Identification of a role for TRIM29 in the control of innate immunity in the respiratory tract
Erratum in
-
Corrigendum: Identification of a role for TRIM29 in the control of innate immunity in the respiratory tract.Nat Immunol. 2016 Nov 16;17(12):1479. doi: 10.1038/ni1216-1479a. Nat Immunol. 2016. PMID: 27849197 No abstract available.
Abstract
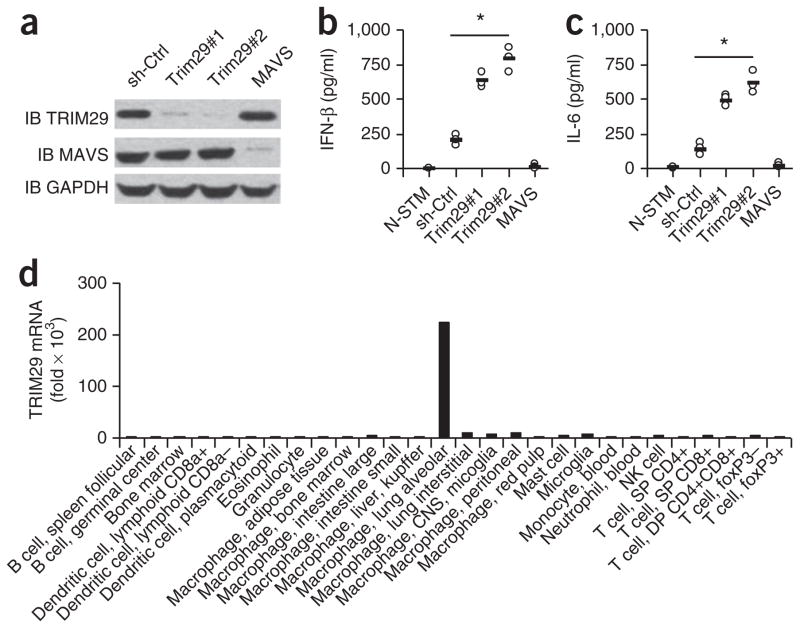
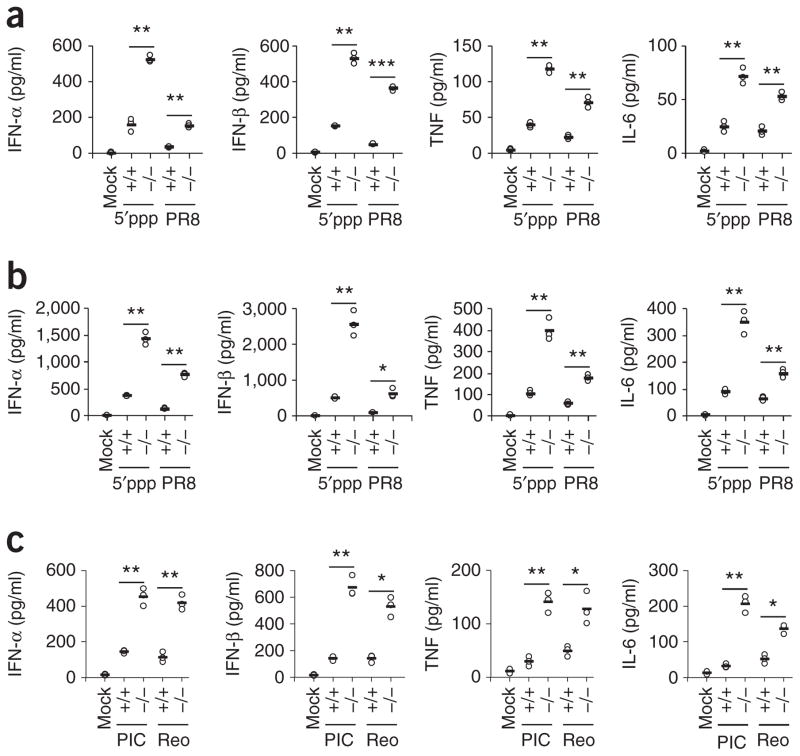
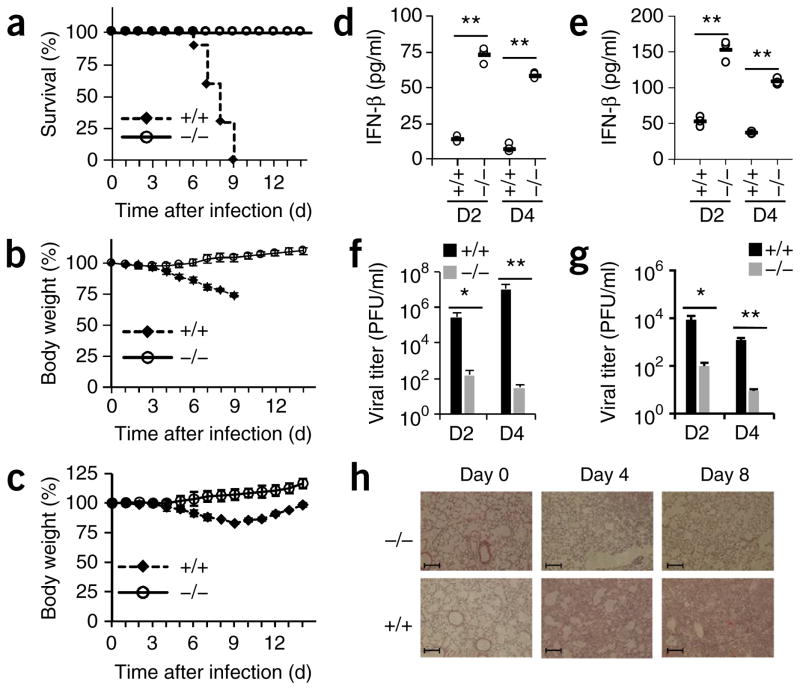
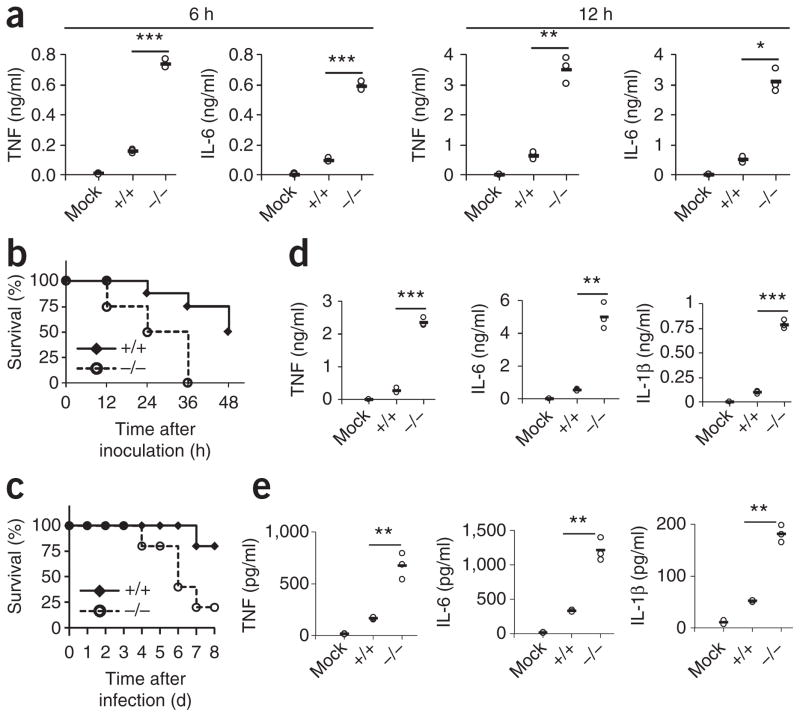
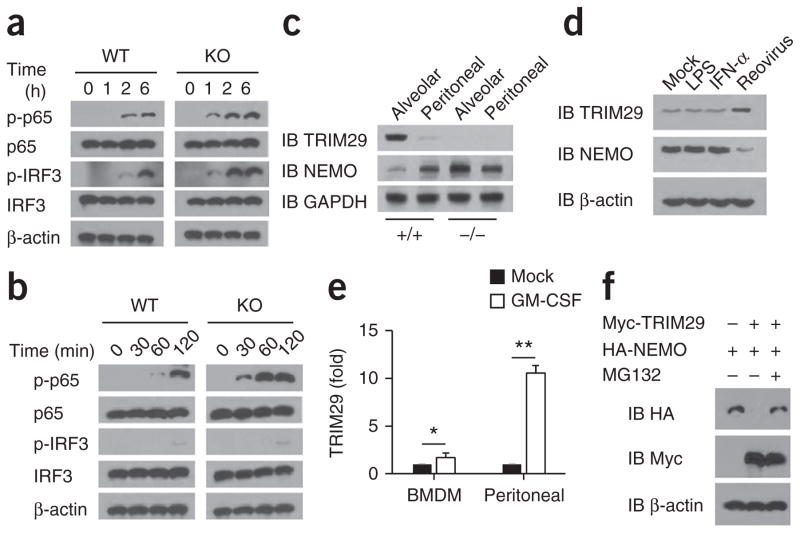
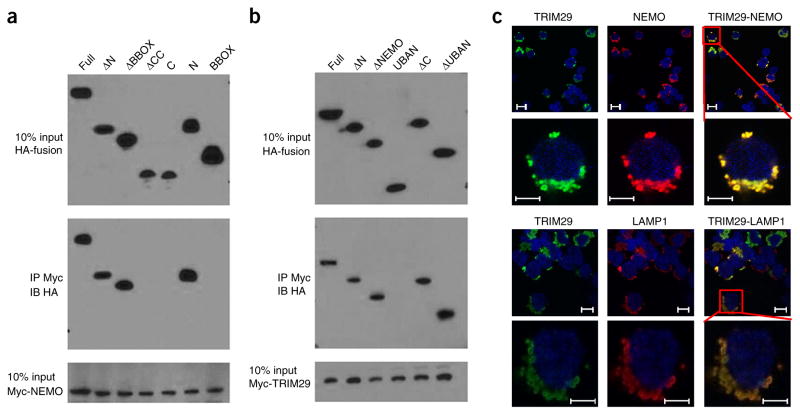
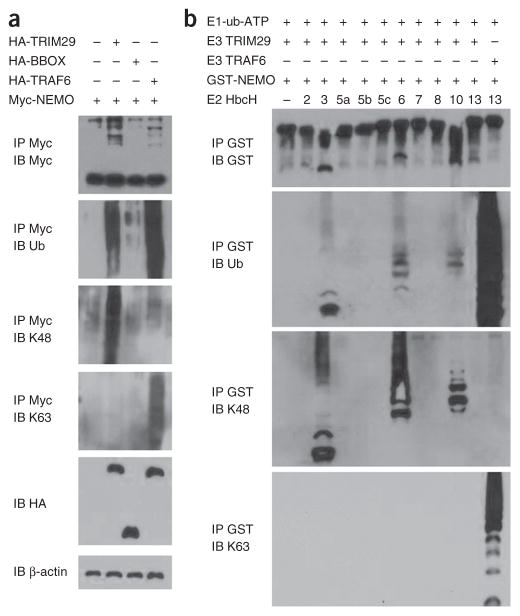
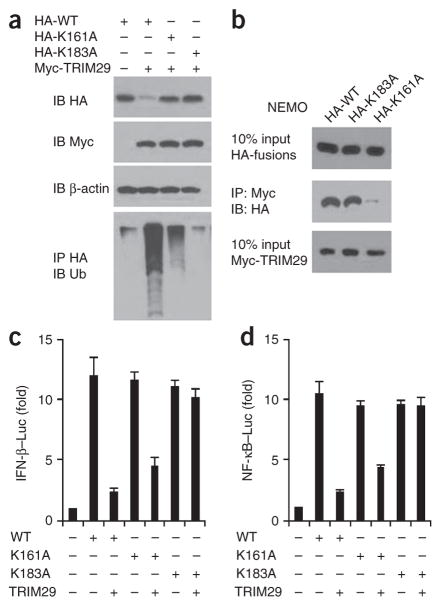
The respiratory tract is heavily populated with innate immune cells, but the mechanisms that control such cells are poorly defined. Here we found that the E3 ubiquitin ligase TRIM29 was a selective regulator of the activation of alveolar macrophages, the expression of type I interferons and the production of proinflammatory cytokines in the lungs. We found that deletion of TRIM29 enhanced macrophage production of type I interferons and protected mice from infection with influenza virus, while challenge of Trim29-/- mice with Haemophilus influenzae resulted in lethal lung inflammation due to massive production of proinflammatory cytokines by macrophages. Mechanistically, we demonstrated that TRIM29 inhibited interferon-regulatory factors and signaling via the transcription factor NF-κB by degrading the adaptor NEMO and that TRIM29 directly bound NEMO and subsequently induced its ubiquitination and proteolytic degradation. These data identify TRIM29 as a key negative regulator of alveolar macrophages and might have important clinical implications for local immunity and immunopathology.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
