

Mycobacteriophages as Incubators for Intein Dissemination and Evolution
- PMID: 27703073
- PMCID: PMC5050341
- DOI: 10.1128/mBio.01537-16
Mycobacteriophages as Incubators for Intein Dissemination and Evolution
Abstract
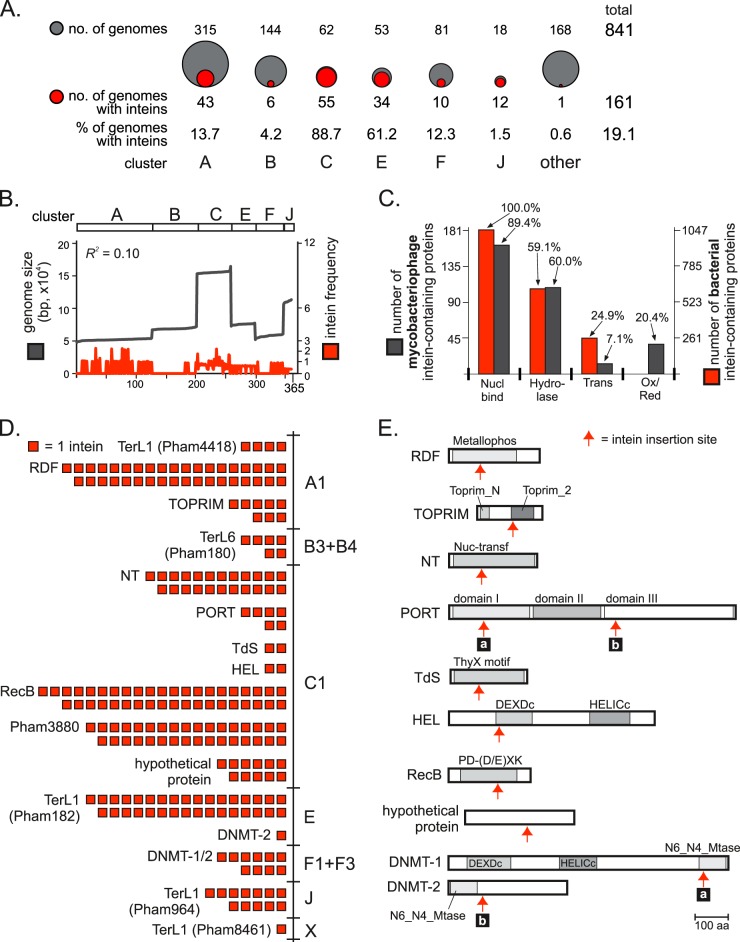
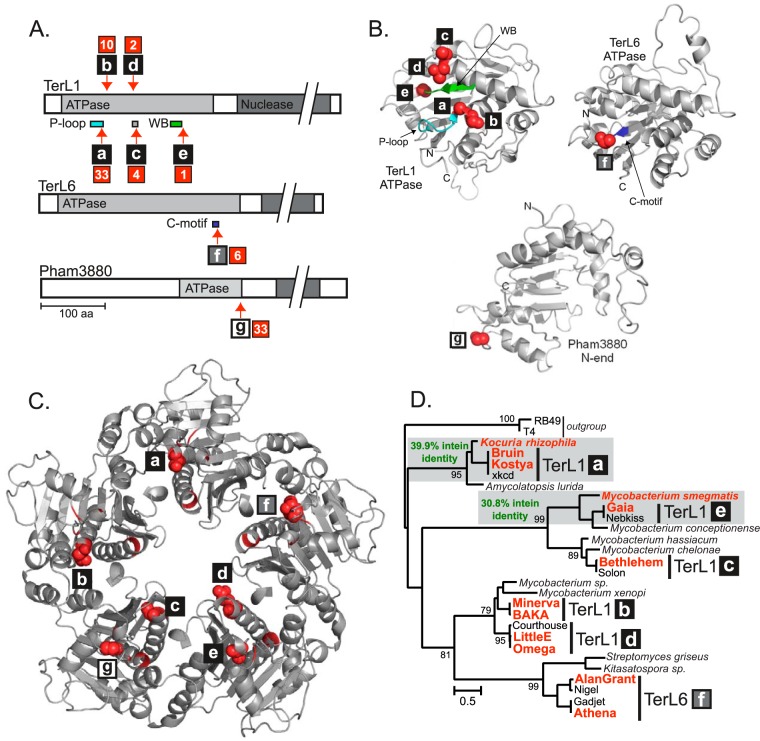
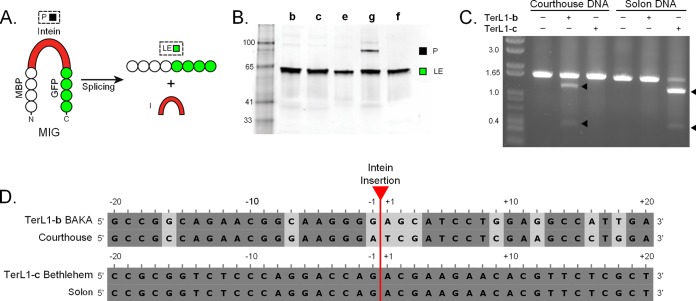
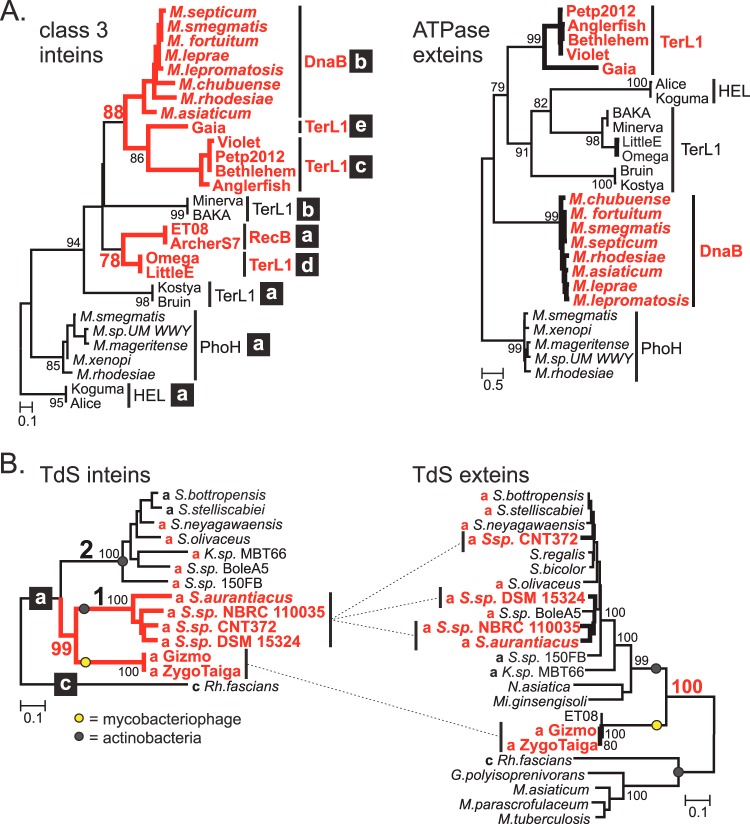
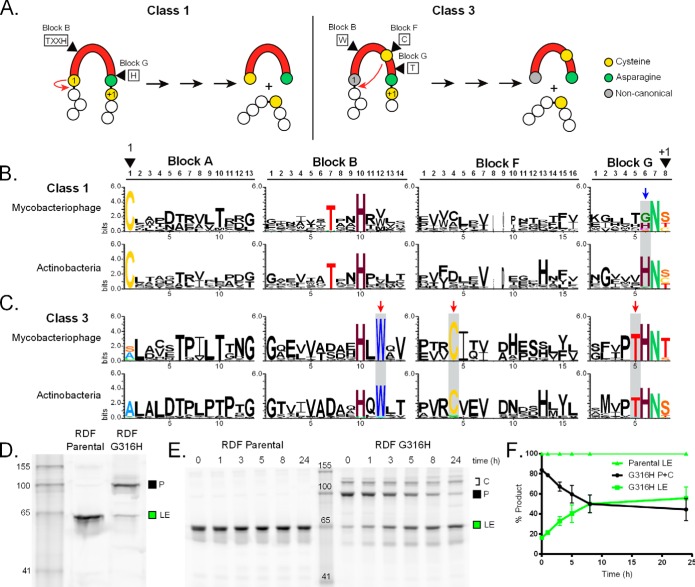
Inteins are self-splicing protein elements that are mobile at the DNA level and are sporadically distributed across microbial genomes. Inteins appear to be horizontally transferred, and it has been speculated that phages may play a role in intein distribution. Our attention turns to mycobacteriophages, which infect mycobacteria, where both phage and host harbor inteins. Using bioinformatics, mycobacteriophage genomes were mined for inteins. This study reveals that these mobile elements are present across multiple mycobacteriophage clusters and are pervasive in certain genes, like the large terminase subunit TerL and a RecB-like nuclease, with the majority of intein-containing genes being phage specific. Strikingly, despite this phage specificity, inteins localize to functional motifs shared with bacteria, such that intein-containing genes have similar roles, like hydrolase activity and nucleic acid binding, indicating a global commonality among intein-hosting proteins. Additionally, there are multiple insertion points within active centers, implying independent invasion events, with regulatory implications. Several phage inteins were shown to be splicing competent and to encode functional homing endonucleases, important for mobility. Further, bioinformatic analysis supports the potential for phages as facilitators of intein movement among mycobacteria and related genera. Analysis of catalytic intein residues finds the highly conserved penultimate histidine inconsistently maintained among mycobacteriophages. Biochemical characterization of a noncanonical phage intein shows that this residue influences precursor accumulation, suggesting that splicing has been tuned in phages to modulate generation of important proteins. Together, this work expands our understanding of phage-based intein dissemination and evolution and implies that phages provide a context for evolution of splicing-based regulation.
Importance: Inteins are mobile protein splicing elements found in critical genes across all domains of life. Mycobacterial inteins are of particular interest because of their occurrence in pathogenic species, such as Mycobacterium tuberculosis and Mycobacterium leprae, which harbor inteins in important proteins. We have discovered a similarity in activities of intein-containing proteins among mycobacteriophages and their intein-rich actinobacterial hosts, with implications for both posttranslational regulation by inteins and phages participating in horizontal intein transfer. Our demonstration of multiple insertion points within active centers of phage proteins implies independent invasion events, indicating the importance of intein maintenance at specific functional sites. The variable conservation of a catalytic splicing residue, leading to profoundly altered splicing rates, points to the regulatory potential of inteins and to mycobacteriophages playing a role in intein evolution. Collectively, these results suggest inteins as posttranslational regulators and mycobacteriophages as both vehicles for intein distribution and incubators for intein evolution.
Copyright © 2016 Kelley et al.
Figures
References
-
- Hirata R, Ohsumk Y, Nakano A, Kawasaki H, Suzuki K, Anraku Y. 1990. Molecular structure of a gene, VMA1, encoding the catalytic subunit of H(+)-translocating adenosine triphosphatase from vacuolar membranes of Saccharomyces cerevisiae. J Biol Chem 265:6726–6733. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
