

A Chemomechanical Model for Nuclear Morphology and Stresses during Cell Transendothelial Migration
- PMID: 27705776
- PMCID: PMC5052451
- DOI: 10.1016/j.bpj.2016.08.011
A Chemomechanical Model for Nuclear Morphology and Stresses during Cell Transendothelial Migration
Abstract
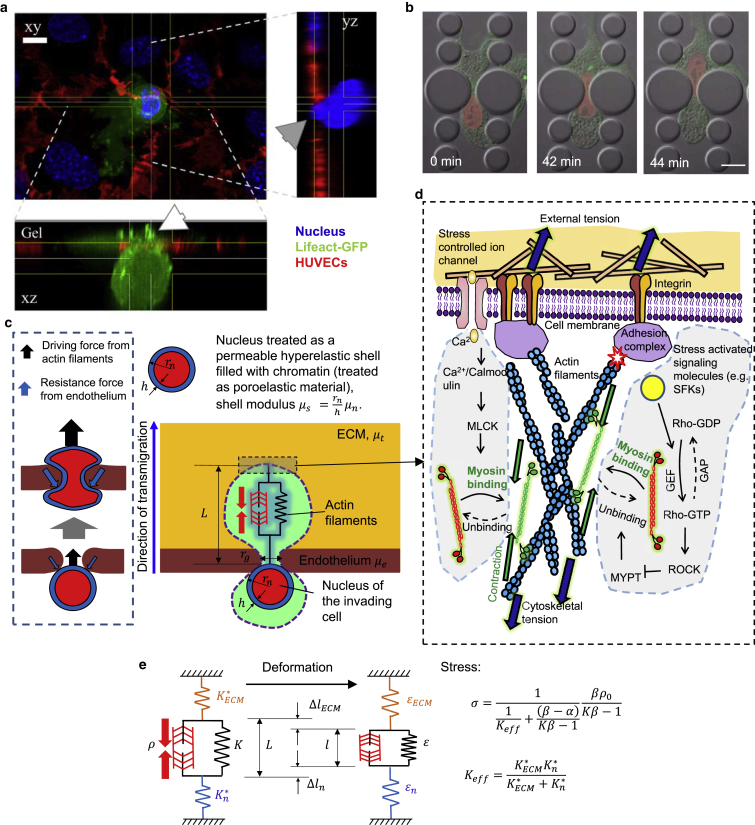
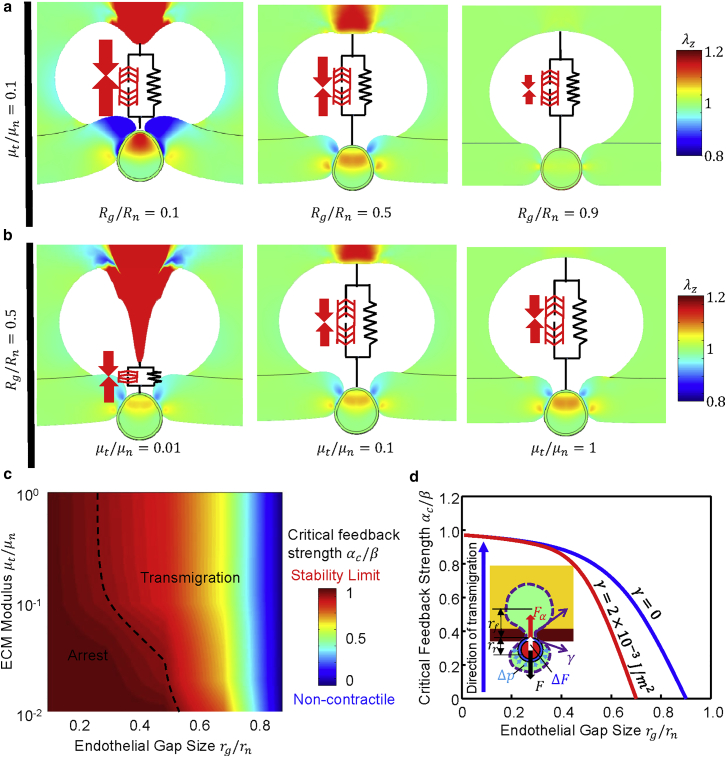
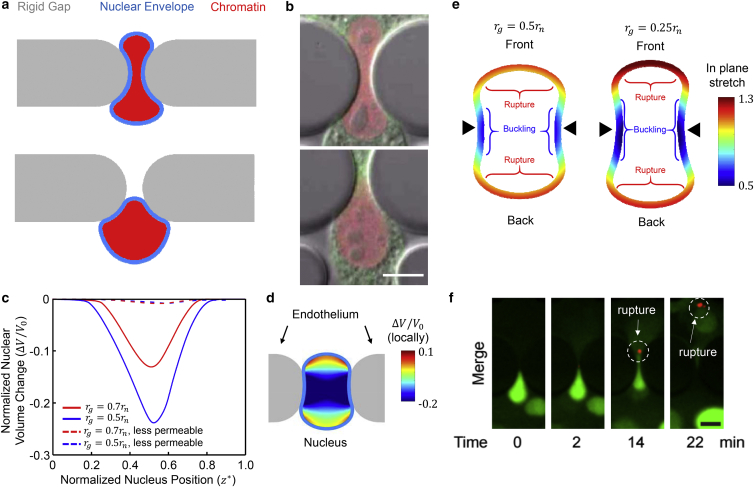
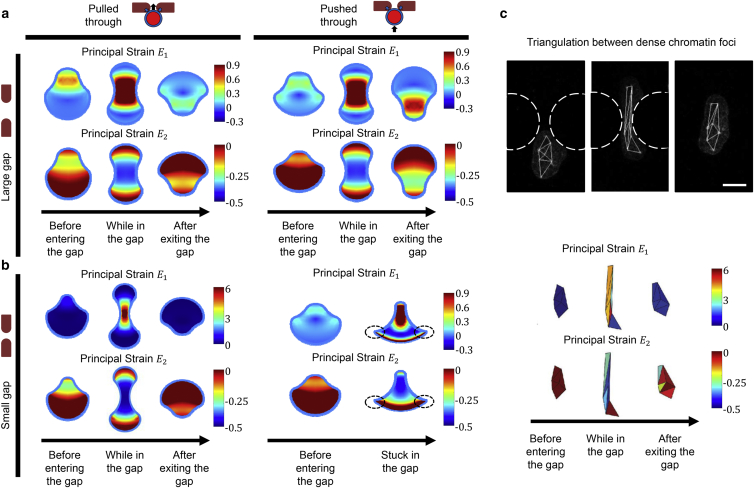
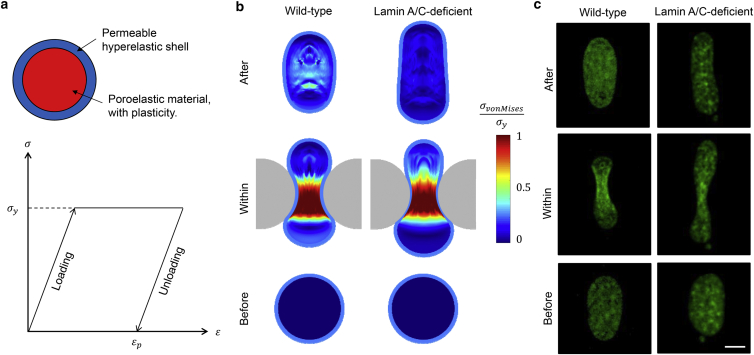
It is now evident that the cell nucleus undergoes dramatic shape changes during important cellular processes such as cell transmigration through extracellular matrix and endothelium. Recent experimental data suggest that during cell transmigration the deformability of the nucleus could be a limiting factor, and the morphological and structural alterations that the nucleus encounters can perturb genomic organization that in turn influences cellular behavior. Despite its importance, a biophysical model that connects the experimentally observed nuclear morphological changes to the underlying biophysical factors during transmigration through small constrictions is still lacking. Here, we developed a universal chemomechanical model that describes nuclear strains and shapes and predicts thresholds for the rupture of the nuclear envelope and for nuclear plastic deformation during transmigration through small constrictions. The model includes actin contraction and cytosolic back pressure that squeeze the nucleus through constrictions and overcome the mechanical resistance from deformation of the nucleus and the constrictions. The nucleus is treated as an elastic shell encompassing a poroelastic material representing the nuclear envelope and inner nucleoplasm, respectively. Tuning the chemomechanical parameters of different components such as cell contractility and nuclear and matrix stiffnesses, our model predicts the lower bounds of constriction size for successful transmigration. Furthermore, treating the chromatin as a plastic material, our model faithfully reproduced the experimentally observed irreversible nuclear deformations after transmigration in lamin-A/C-deficient cells, whereas the wild-type cells show much less plastic deformation. Along with making testable predictions, which are in accord with our experiments and existing literature, our work provides a realistic framework to assess the biophysical modulators of nuclear deformation during cell transmigration.
Copyright © 2016 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
