

Strict in vivo specificity of the Bcl11a erythroid enhancer
- PMID: 27707736
- PMCID: PMC5106112
- DOI: 10.1182/blood-2016-08-736249
Strict in vivo specificity of the Bcl11a erythroid enhancer
Abstract
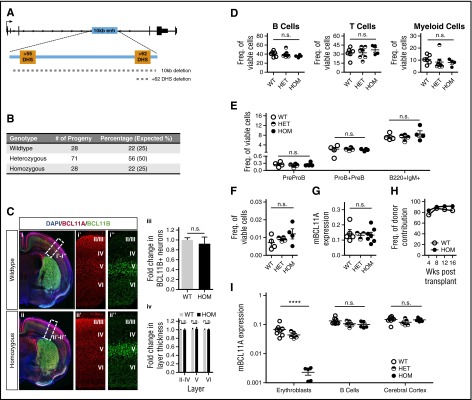
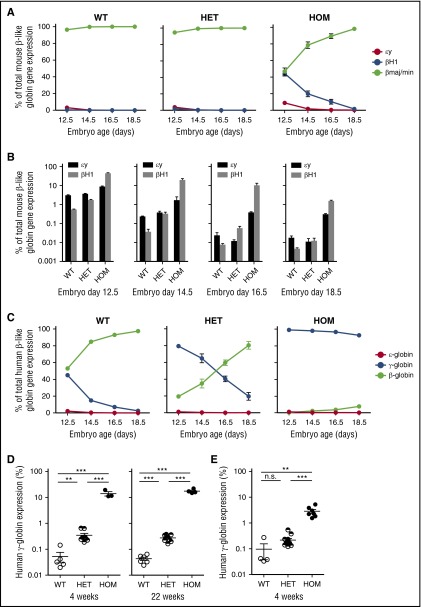
BCL11A, a repressor of human fetal (γ-)globin expression, is required for immune and hematopoietic stem cell functions and brain development. Regulatory sequences within the gene, which are subject to genetic variation affecting fetal globin expression, display hallmarks of an erythroid enhancer in cell lines and transgenic mice. As such, this enhancer is a novel, attractive target for therapeutic gene editing. To explore the roles of such sequences in vivo, we generated mice in which the orthologous 10-kb intronic sequences were removed. Bcl11a enhancer-deleted mice, Bcl11a(Δenh), phenocopy the BCL11A-null state with respect to alterations of globin expression, yet are viable and exhibit no observable blood, brain, or other abnormalities. These preclinical findings provide strong in vivo support for genetic modification of the enhancer for therapy of hemoglobin disorders.
© 2016 by The American Society of Hematology.
Figures
References
-
- Menzel S, Garner C, Gut I, et al. A QTL influencing F cell production maps to a gene encoding a zinc-finger protein on chromosome 2p15. Nat Genet. 2007;39(10):1197–1199. - PubMed
-
- Sankaran VG, Menne TF, Xu J, et al. Human fetal hemoglobin expression is regulated by the developmental stage-specific repressor BCL11A. Science. 2008;322(5909):1839–1842. - PubMed
MeSH terms
Substances
Grants and funding
- R56 NS093376/NS/NINDS NIH HHS/United States
- K08 DK093705/DK/NIDDK NIH HHS/United States
- R37 NS041590/NS/NINDS NIH HHS/United States
- R01 HL032259/HL/NHLBI NIH HHS/United States
- R03 DK109232/DK/NIDDK NIH HHS/United States
- R01 NS041590/NS/NINDS NIH HHS/United States
- P30 DK049216/DK/NIDDK NIH HHS/United States
- F31 NS064730/NS/NINDS NIH HHS/United States
- T32 GM007753/GM/NIGMS NIH HHS/United States
- P01 HL032262/HL/NHLBI NIH HHS/United States
- R01 NS045523/NS/NINDS NIH HHS/United States
- U54 DK110805/DK/NIDDK NIH HHS/United States
- F31 NS080343/NS/NINDS NIH HHS/United States
- R01 NS075672/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
