

The Tetherin Antagonism of the Ebola Virus Glycoprotein Requires an Intact Receptor-Binding Domain and Can Be Blocked by GP1-Specific Antibodies
- PMID: 27707924
- PMCID: PMC5126366
- DOI: 10.1128/JVI.01563-16
The Tetherin Antagonism of the Ebola Virus Glycoprotein Requires an Intact Receptor-Binding Domain and Can Be Blocked by GP1-Specific Antibodies
Abstract
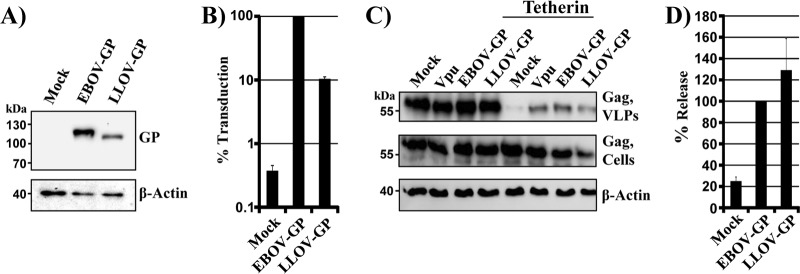
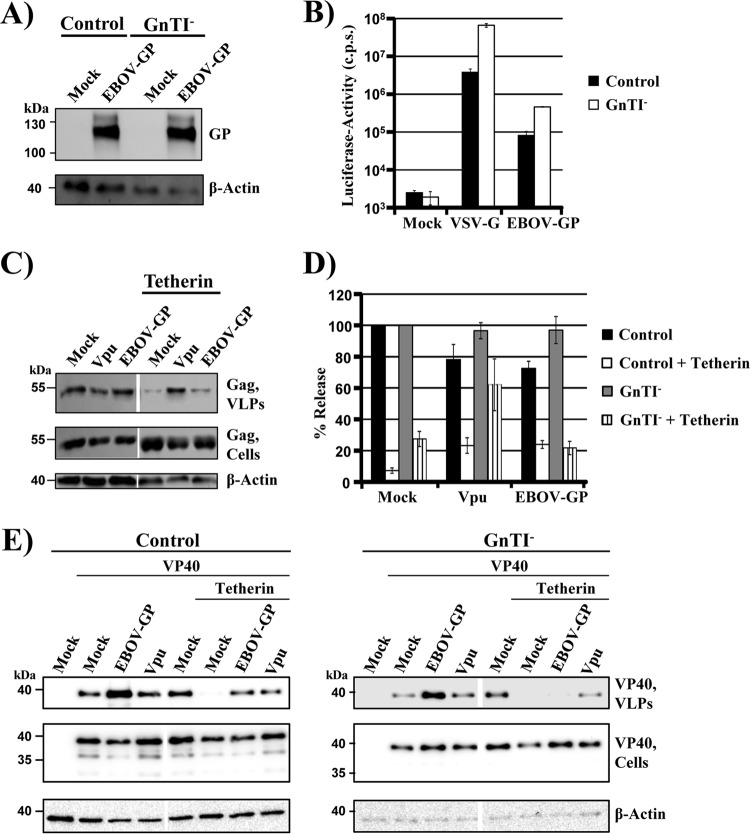
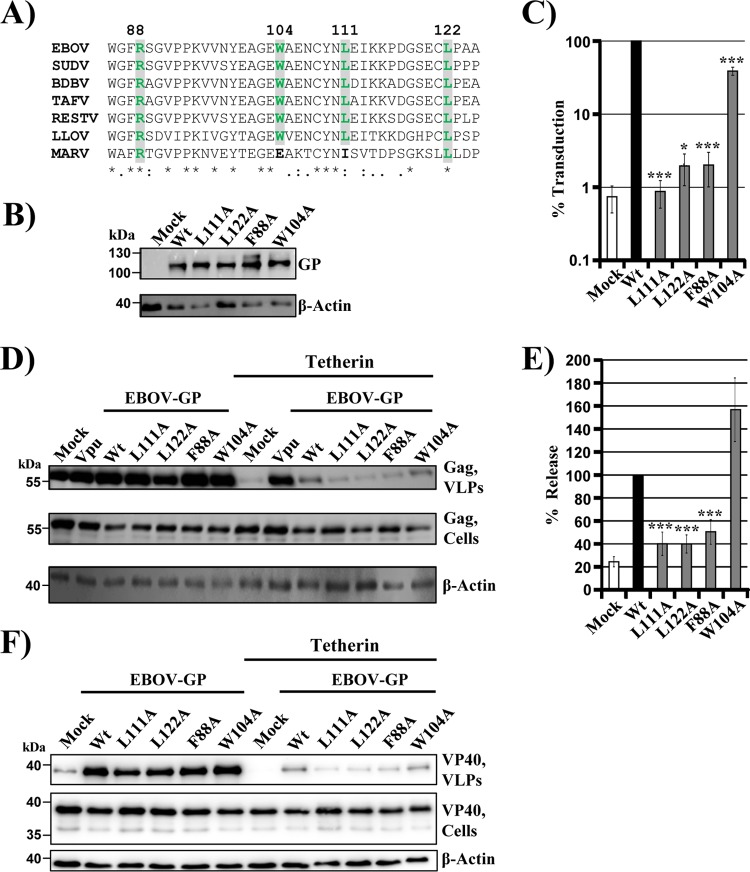
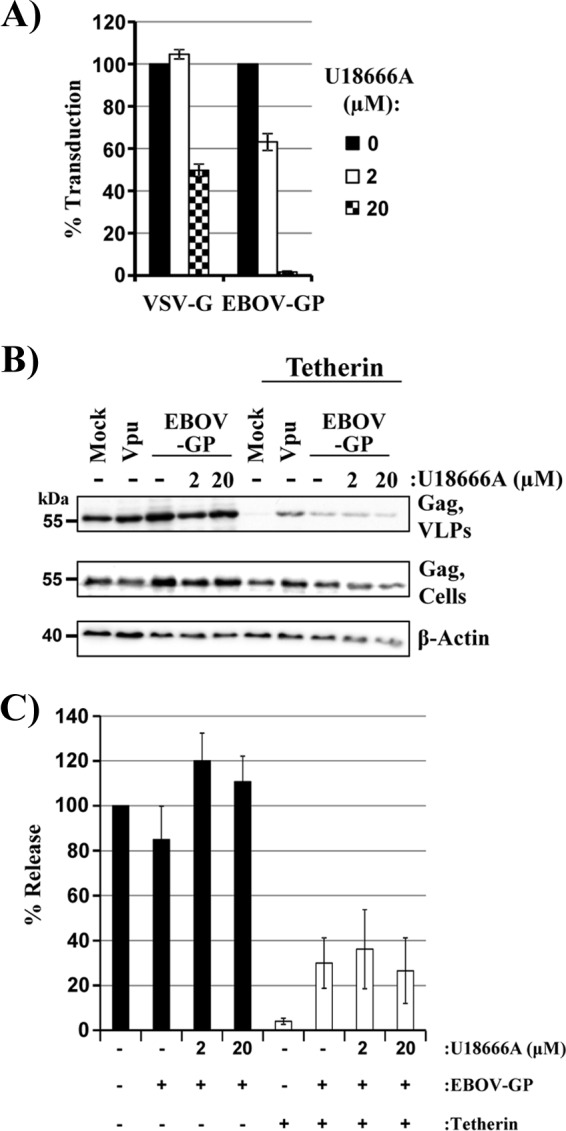
The glycoprotein of Ebola virus (EBOV GP), a member of the family Filoviridae, facilitates viral entry into target cells. In addition, EBOV GP antagonizes the antiviral activity of the host cell protein tetherin, which may otherwise restrict EBOV release from infected cells. However, it is unclear how EBOV GP antagonizes tetherin, and it is unknown whether the GP of Lloviu virus (LLOV), a filovirus found in dead bats in Northern Spain, also counteracts tetherin. Here, we show that LLOV GP antagonizes tetherin, indicating that tetherin may not impede LLOV spread in human cells. Moreover, we demonstrate that appropriate processing of N-glycans in tetherin/GP-coexpressing cells is required for tetherin counteraction by EBOV GP. Furthermore, we show that an intact receptor-binding domain (RBD) in the GP1 subunit of EBOV GP is a prerequisite for tetherin counteraction. In contrast, blockade of Niemann-Pick disease type C1 (NPC1), a cellular binding partner of the RBD, did not interfere with tetherin antagonism. Finally, we provide evidence that an antibody directed against GP1, which protects mice from a lethal EBOV challenge, may block GP-dependent tetherin antagonism. Our data, in conjunction with previous reports, indicate that tetherin antagonism is conserved among the GPs of all known filoviruses and demonstrate that the GP1 subunit of EBOV GP plays a central role in tetherin antagonism.
Importance: Filoviruses are reemerging pathogens that constitute a public health threat. Understanding how Ebola virus (EBOV), a highly pathogenic filovirus responsible for the 2013-2016 Ebola virus disease epidemic in western Africa, counteracts antiviral effectors of the innate immune system might help to define novel targets for antiviral intervention. Similarly, determining whether Lloviu virus (LLOV), a filovirus detected in bats in northern Spain, is inhibited by innate antiviral effectors in human cells might help to determine whether the virus constitutes a threat to humans. The present study shows that LLOV, like EBOV, counteracts the antiviral effector protein tetherin via its glycoprotein (GP), suggesting that tetherin does not pose a defense against LLOV spread in humans. Moreover, our work identifies the GP1 subunit of EBOV GP, in particular an intact receptor-binding domain, as critical for tetherin counteraction and provides evidence that antibodies directed against GP1 can interfere with tetherin counteraction.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- WHO. 17 February 2016. Ebola situation report-17 February 2016. http://apps.who.int/ebola/current-situation/ebola-situation-report-17-fe....
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
