

Phosphorylation Requirement of Murine Leukemia Virus p12
- PMID: 27707931
- PMCID: PMC5126377
- DOI: 10.1128/JVI.01178-16
Phosphorylation Requirement of Murine Leukemia Virus p12
Abstract
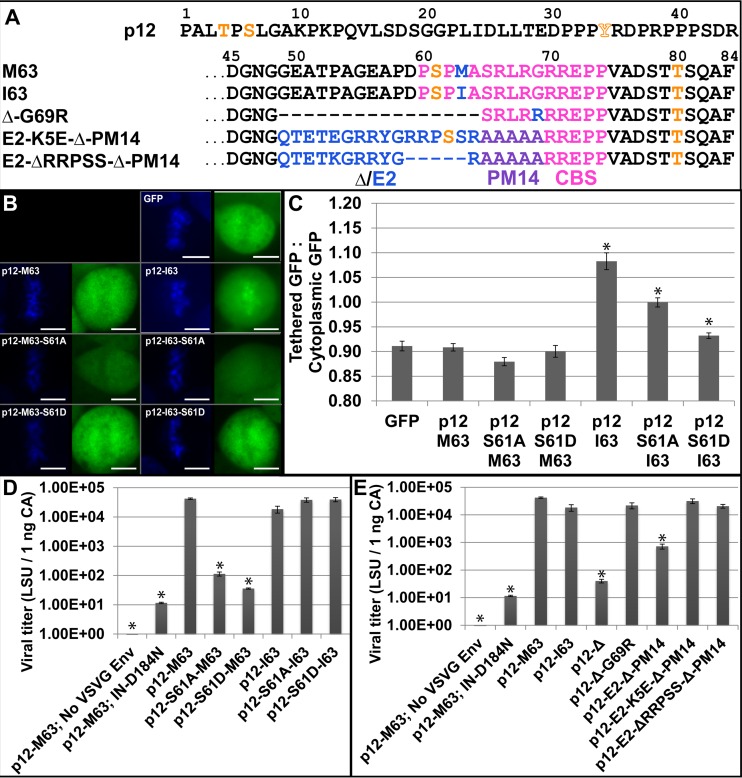
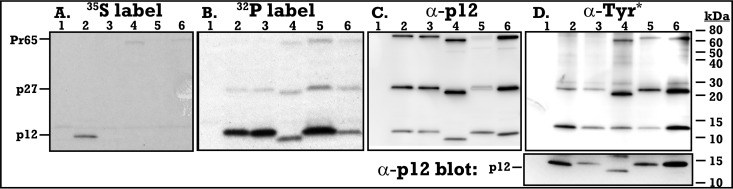
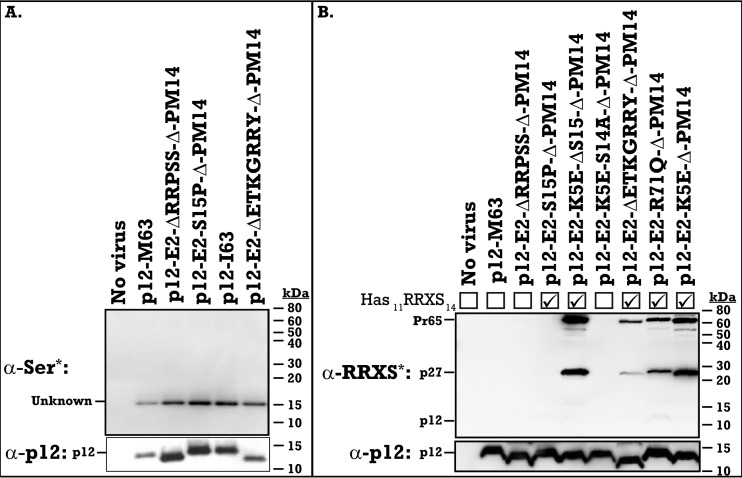
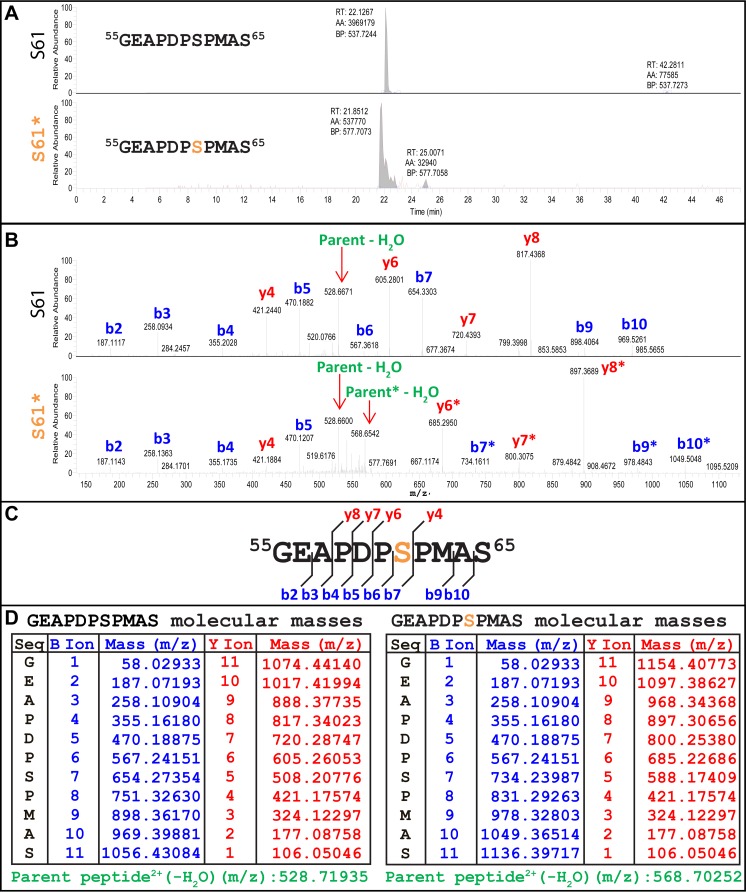
The p12 protein of murine leukemia virus (MLV) Gag is associated with the preintegration complex (PIC), and mutants of p12 (PM14) exhibit defects in nuclear entry/retention. Mutants of the phosphorylated serine 61 also have been reported to have defects in the early life cycle. Here we show that a phosphorylated peptide motif derived from human papillomavirus 8 (HPV-8), the E2 hinge region including residues 240 to 255, can functionally replace the main phosphorylated motif of MLV p12 and can rescue the viral titer of a strain with the lethal p12-PM14 mutation. Complementation with the HPV-8 E2 hinge motif generated multiple second-site mutations in live viral passage assays. Additional p12 phosphorylation sites were detected, including the late domain of p12 (PPPY) as well as the late domain/protease cleavage site of matrix (LYPAL), by mass spectrometry and Western blotting. Chromatin binding of p12-green fluorescent protein (GFP) fusion protein and functional complementation of p12-PM14 occurred in a manner independent of the E2 hinge region phosphorylation. Replacement of serine 61 by alanine within the minimal tethering domain (61SPMASRLRGRR71) maintained tethering, but in the context of the full-length p12, mutants with substitutions in S61 remained untethered and lost infectivity, indicating phosphorylation of p12 serine 61 functions to temporally regulate early and late p12 functions.
Importance: The p12 protein, required for both early and late viral functions, is the predominant phosphorylated viral protein of Moloney MLV and is required for virus viability. Our studies indicate that the N terminus of p12 represses the early function of the chromatin binding domain and that deletion of the N terminus activates chromatin binding in the wild-type Moloney MLV p12 protein. Mass spectrometry and mutagenesis studies suggest that phosphorylation of both the repression domain and the chromatin binding domain acts to temporally regulate this process at the appropriate stages during infection.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Repression of the Chromatin-Tethering Domain of Murine Leukemia Virus p12.J Virol. 2016 Nov 28;90(24):11197-11207. doi: 10.1128/JVI.01084-16. Print 2016 Dec 15. J Virol. 2016. PMID: 27707926 Free PMC article.
-
Murine leukemia virus p12 tethers the capsid-containing pre-integration complex to chromatin by binding directly to host nucleosomes in mitosis.PLoS Pathog. 2018 Jun 15;14(6):e1007117. doi: 10.1371/journal.ppat.1007117. eCollection 2018 Jun. PLoS Pathog. 2018. PMID: 29906285 Free PMC article.
-
Infectivity of Moloney murine leukemia virus defective in late assembly events is restored by late assembly domains of other retroviruses.J Virol. 2000 Aug;74(16):7250-60. doi: 10.1128/jvi.74.16.7250-7260.2000. J Virol. 2000. PMID: 10906179 Free PMC article.
-
Phosphorylated serine residues and an arginine-rich domain of the moloney murine leukemia virus p12 protein are required for early events of viral infection.J Virol. 2003 Feb;77(3):1820-9. doi: 10.1128/jvi.77.3.1820-1829.2003. J Virol. 2003. PMID: 12525616 Free PMC article.
-
Destroying retroviruses from within.Trends Microbiol. 1996 Nov;4(11):421-6. doi: 10.1016/0966-842x(96)10065-2. Trends Microbiol. 1996. PMID: 8950810 Review.
Cited by
-
Repression of the Chromatin-Tethering Domain of Murine Leukemia Virus p12.J Virol. 2016 Nov 28;90(24):11197-11207. doi: 10.1128/JVI.01084-16. Print 2016 Dec 15. J Virol. 2016. PMID: 27707926 Free PMC article.
-
Evidence for Tethering of Human Cytomegalovirus Genomes to Host Chromosomes.Front Cell Infect Microbiol. 2020 Sep 30;10:577428. doi: 10.3389/fcimb.2020.577428. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33117732 Free PMC article.
-
Post-mitotic BET-induced reshaping of integrase quaternary structure supports wild-type MLV integration.Nucleic Acids Res. 2019 Feb 20;47(3):1195-1210. doi: 10.1093/nar/gky1157. Nucleic Acids Res. 2019. PMID: 30445610 Free PMC article.
-
Targeting the Nucleosome Acidic Patch by Viral Proteins: Two Birds with One Stone?mBio. 2022 Apr 26;13(2):e0173321. doi: 10.1128/mbio.01733-21. Epub 2022 Mar 28. mBio. 2022. PMID: 35343785 Free PMC article. Review.
-
BET-Independent Murine Leukemia Virus Integration Is Retargeted In Vivo and Selects Distinct Genomic Elements for Lymphomagenesis.Microbiol Spectr. 2022 Aug 31;10(4):e0147822. doi: 10.1128/spectrum.01478-22. Epub 2022 Jul 19. Microbiol Spectr. 2022. PMID: 35852337 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
