

ERK/c-Jun Recruits Tet1 to Induce Zta Expression and Epstein-Barr Virus Reactivation through DNA Demethylation
- PMID: 27708396
- PMCID: PMC5052586
- DOI: 10.1038/srep34543
ERK/c-Jun Recruits Tet1 to Induce Zta Expression and Epstein-Barr Virus Reactivation through DNA Demethylation
Abstract
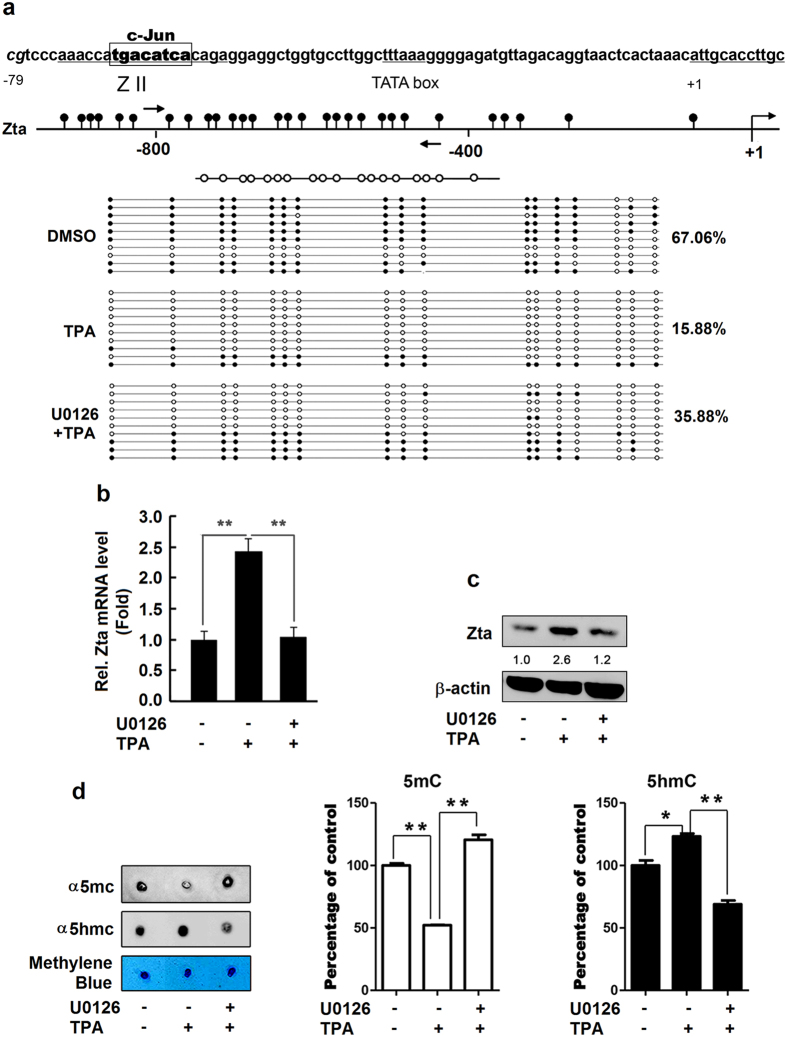
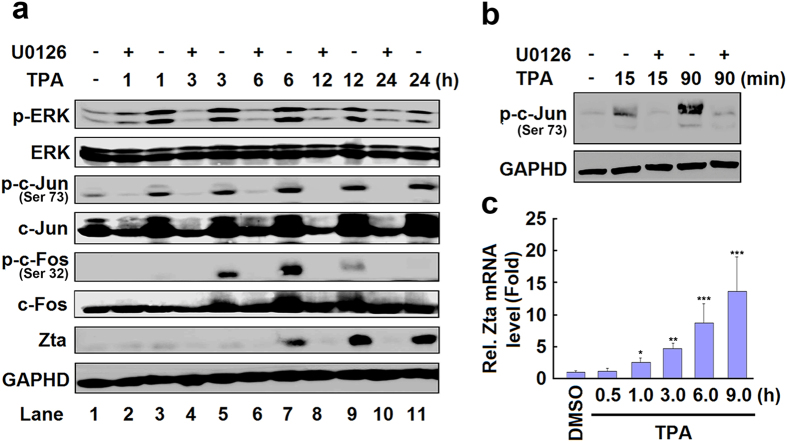
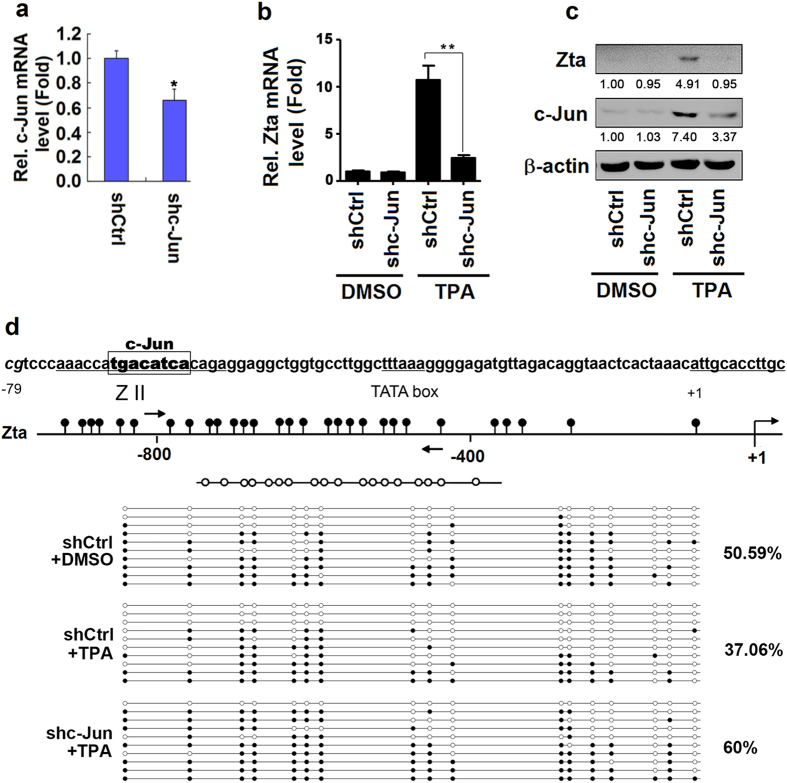
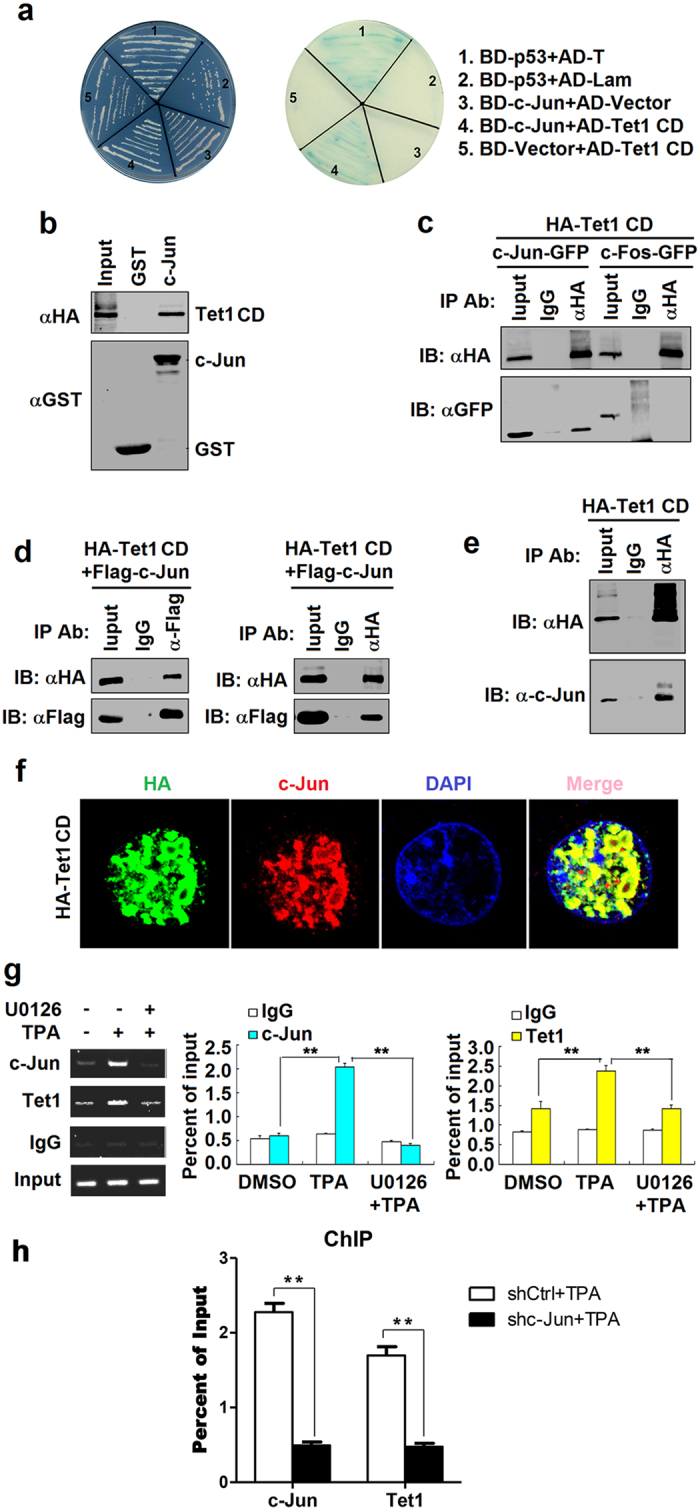
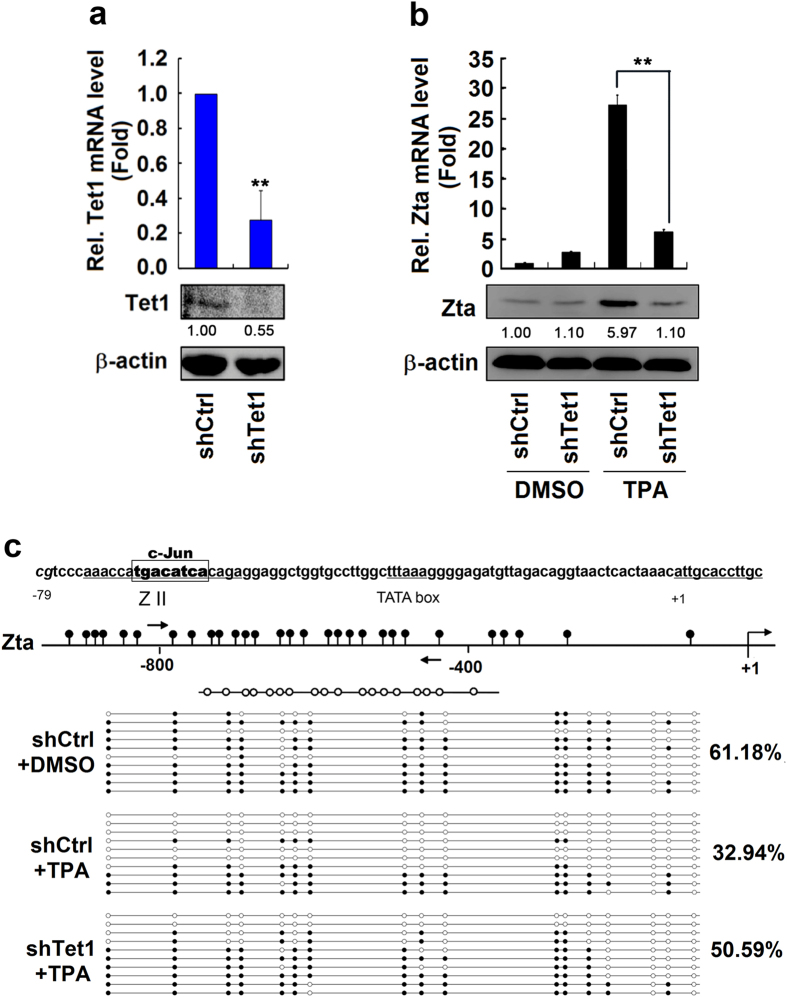
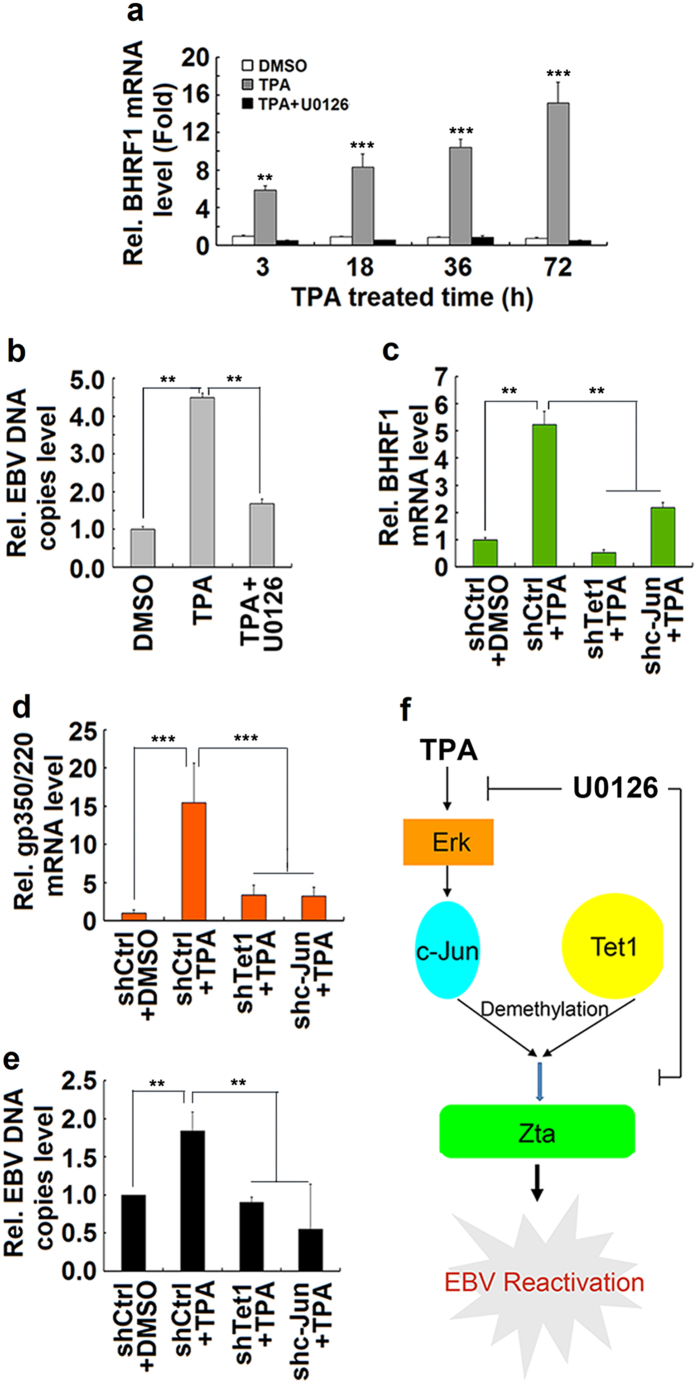
DNA demethylation plays an essential role in the reactivation of Epstein-Barr virus (EBV) from latency infection. However, it is unclear how epigenetic modification is initiated in responding to stimuli. Here, we demonstrate that ERK/c-Jun signaling is involved in DNA demethylation of EBV immediate early (IE) gene Zta in response to 12-O-Tetradecanoylphorbol-13-acetate (TPA) stimulation. Remarkably, Ser73 phosphorylation of c-Jun facilitates Zta promoter demethylation and EBV reactivation, whereas knockdown of c-Jun attenuates Zta demethylation and viral reactivation. More importantly, we reveal for the first time that c-Jun interacts with DNA dioxygenase Tet1 and facilitates Tet1 to bind to Zta promoter. The binding of c-Jun and Tet1 to Zta enhances promoter demethylation, resulting in the activation of Zta, the stimulation of BHRF1 (a lytic early gene) and gp350/220 (a lytic late gene), and ultimately the reactivation of EBV. Knockdown of Tet1 attenuates TPA-induced Zta demethylation and EBV reactivation. Thus, TPA activates ERK/c-Jun signaling, which subsequently facilitates Tet1 to bind to Zta promoter, leading to DNA demethylation, gene expression, and EBV reactivation. This study reveals important roles of ERK/c-Jun signaling and Tet1 dioxygenase in epigenetic modification, and provides new insights into the mechanism underlying the regulation of virus latent and lytic infection.
Figures
References
-
- Salvetti M., Giovannoni G. & Aloisi F. Epstein-Barr virus and multiple sclerosis. Curr Opin Neurol 22, 201–206 (2009). - PubMed
-
- Young L. S. & Rickinson A. B. Epstein-Barr virus: 40 years on. Nat Rev Cancer 4, 757–768 (2004). - PubMed
-
- Epstein M. A., Achong B. G. & Barr Y. M. Virus Particles in Cultured Lymphoblasts from Burkitt’s Lymphoma. Lancet 1, 702–703 (1964). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
