

Construction of Asymmetrical Hexameric Biomimetic Motors with Continuous Single-Directional Motion by Sequential Coordination
- PMID: 27709780
- PMCID: PMC5217803
- DOI: 10.1002/smll.201601600
Construction of Asymmetrical Hexameric Biomimetic Motors with Continuous Single-Directional Motion by Sequential Coordination
Abstract
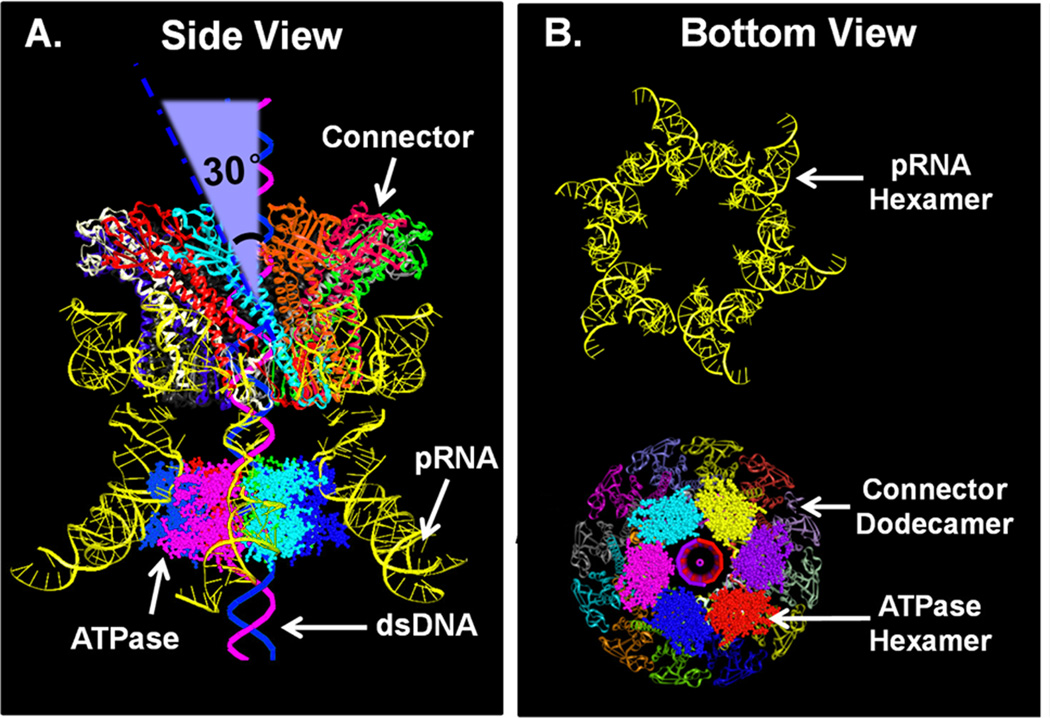
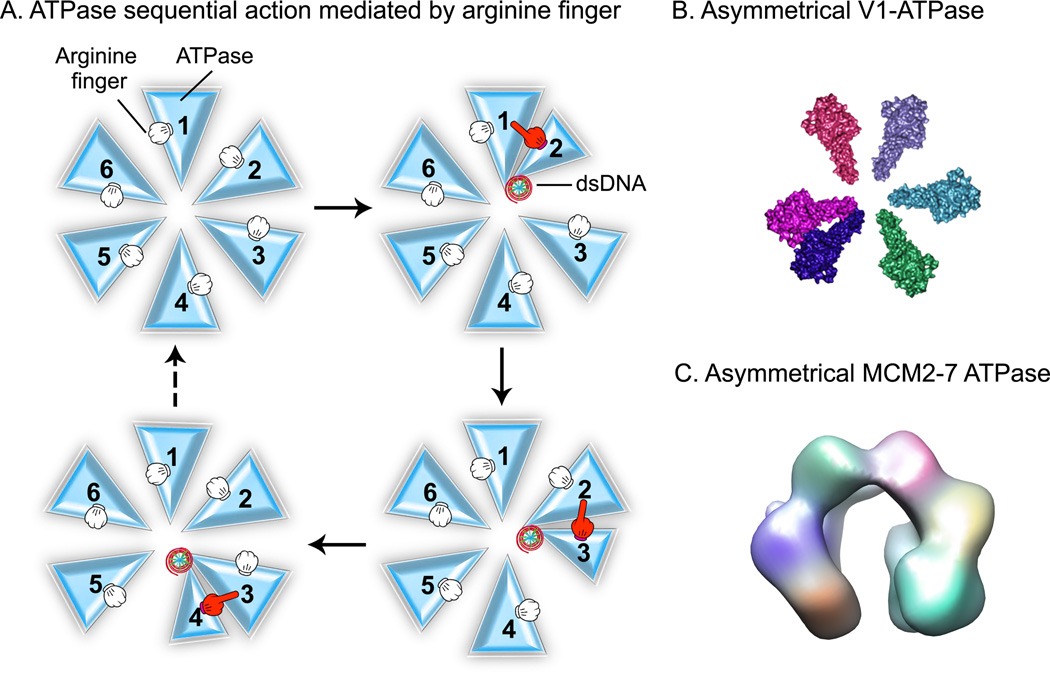
The significance of bionanomotors in nanotechnology is analogous to mechanical motors in daily life. Here the principle and approach for designing and constructing biomimetic nanomotors with continuous single-directional motion are reported. This bionanomotor is composed of a dodecameric protein channel, a six-pRNA ring, and an ATPase hexamer. Based on recent elucidations of the one-way revolving mechanisms of the phi29 double-stranded DNA (dsDNA) motor, various RNA and protein elements are designed and tested by single-molecule imaging and biochemical assays, with which the motor with active components has been constructed. The motor motion direction is controlled by three operation elements: (1) Asymmetrical ATPase with ATP-interacting domains for alternative DNA binding/pushing regulated by an arginine finger in a sequential action manner. The arginine finger bridges two adjacent ATPase subunits into a non-covalent dimer, resulting in an asymmetrical hexameric complex containing one dimer and four monomers. (2) The dsDNA translocation channel as a one-way valve. (3) The hexameric pRNA ring geared with left-/right-handed loops. Assessments of these constructs reveal that one inactive subunit of pRNA/ATPase is sufficient to completely block motor function (defined as K = 1), implying that these components work sequentially based on the principle of binomial distribution and Yang Hui's triangle.
Keywords: bionanomotors; inter-subunit communication; one-way traffic; real-time recording; single-molecule imaging.
© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures






Similar articles
-
Revolving ATPase motors as asymmetrical hexamers in translocating lengthy dsDNA via conformational changes and electrostatic interactions in phi29, T7, herpesvirus, mimivirus, E. coli, and Streptomyces.Exploration (Beijing). 2023 Feb 5;3(2):20210056. doi: 10.1002/EXP.20210056. eCollection 2023 Apr. Exploration (Beijing). 2023. PMID: 37324034 Free PMC article. Review.
-
An Arginine Finger Regulates the Sequential Action of Asymmetrical Hexameric ATPase in the Double-Stranded DNA Translocation Motor.Mol Cell Biol. 2016 Sep 12;36(19):2514-23. doi: 10.1128/MCB.00142-16. Print 2016 Oct 1. Mol Cell Biol. 2016. PMID: 27457616 Free PMC article.
-
Identification of Arginine Finger as the Starter of the Biomimetic Motor in Driving Double-Stranded DNA.ACS Nano. 2021 Aug 24;15(8):13260-13266. doi: 10.1021/acsnano.1c02973. Epub 2021 Jul 29. ACS Nano. 2021. PMID: 34323470 Free PMC article.
-
Common mechanisms of DNA translocation motors in bacteria and viruses using one-way revolution mechanism without rotation.Biotechnol Adv. 2014 Jul-Aug;32(4):853-72. doi: 10.1016/j.biotechadv.2014.01.006. Biotechnol Adv. 2014. PMID: 24913057 Free PMC article. Review.
-
Ultrastable pRNA hexameric ring gearing hexameric phi29 DNA-packaging motor by revolving without rotating and coiling.Curr Opin Biotechnol. 2013 Aug;24(4):581-90. doi: 10.1016/j.copbio.2013.03.019. Epub 2013 May 14. Curr Opin Biotechnol. 2013. PMID: 23683853 Free PMC article. Review.
Cited by
-
Structural Dynamics of Nonenveloped Virus Disassembly Intermediates.J Virol. 2019 Oct 29;93(22):e01115-19. doi: 10.1128/JVI.01115-19. Print 2019 Nov 15. J Virol. 2019. PMID: 31484752 Free PMC article.
-
Revolving ATPase motors as asymmetrical hexamers in translocating lengthy dsDNA via conformational changes and electrostatic interactions in phi29, T7, herpesvirus, mimivirus, E. coli, and Streptomyces.Exploration (Beijing). 2023 Feb 5;3(2):20210056. doi: 10.1002/EXP.20210056. eCollection 2023 Apr. Exploration (Beijing). 2023. PMID: 37324034 Free PMC article. Review.
-
DNA/RNA hybrids avoid channel gating that leads to the continued packaging of numerous hybrids into the phi29 protein shell.Nucleic Acids Res. 2025 Mar 20;53(6):gkaf242. doi: 10.1093/nar/gkaf242. Nucleic Acids Res. 2025. PMID: 40193705 Free PMC article.
-
Controlling the Revolving and Rotating Motion Direction of Asymmetric Hexameric Nanomotor by Arginine Finger and Channel Chirality.ACS Nano. 2019 Jun 25;13(6):6207-6223. doi: 10.1021/acsnano.8b08849. Epub 2019 May 28. ACS Nano. 2019. PMID: 31067030 Free PMC article. Review.
-
Revolving hexameric ATPases as asymmetric motors to translocate double-stranded DNA genome along one strand.iScience. 2023 May 19;26(6):106922. doi: 10.1016/j.isci.2023.106922. eCollection 2023 Jun 16. iScience. 2023. PMID: 37305704 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
