

Scaffold Role of DUSP22 in ASK1-MKK7-JNK Signaling Pathway
- PMID: 27711255
- PMCID: PMC5053508
- DOI: 10.1371/journal.pone.0164259
Scaffold Role of DUSP22 in ASK1-MKK7-JNK Signaling Pathway
Abstract
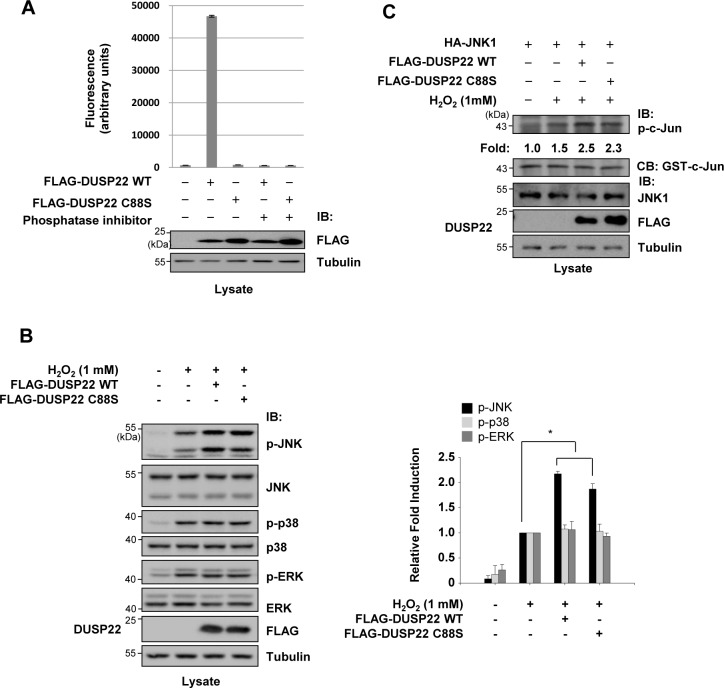
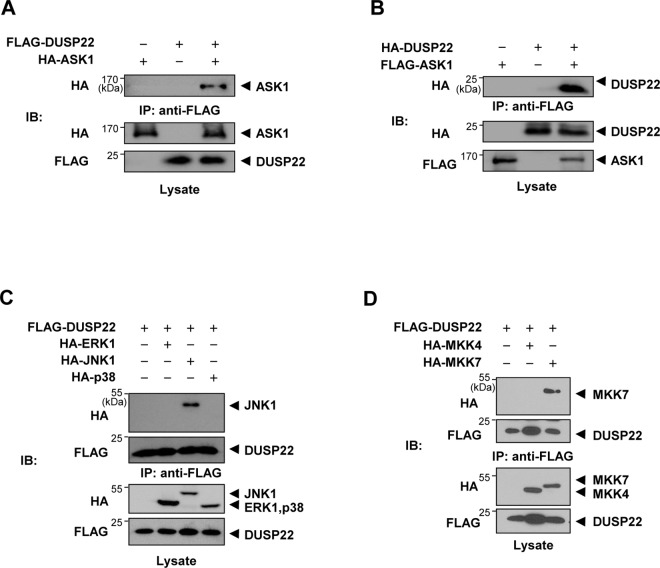
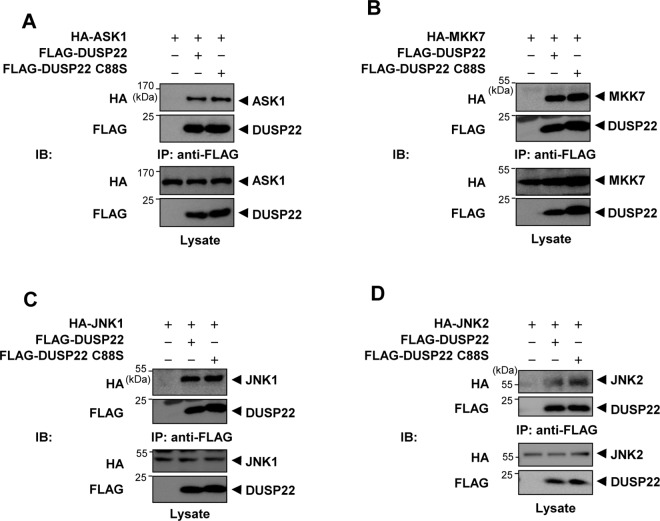
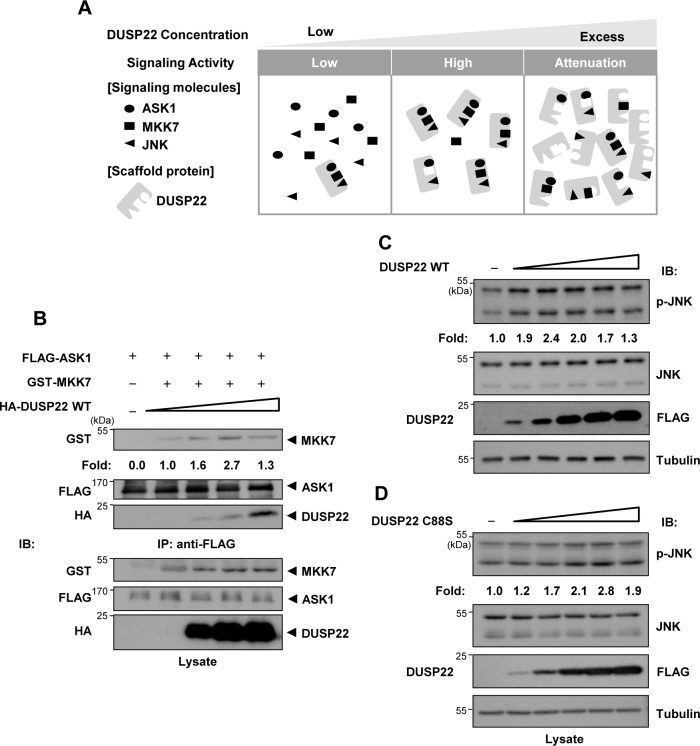
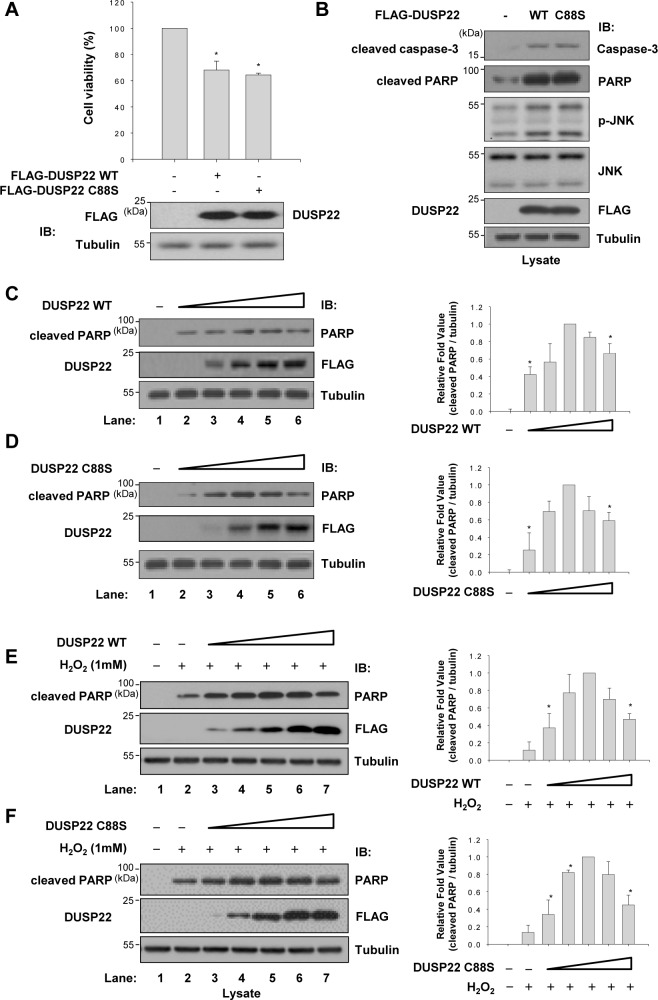
Mitogen-activated protein kinases (MAPKs) are involved in a variety of intracellular events such as gene expression, cell proliferation, and programmed cell death. MAPKs are activated by dual phosphorylation on threonine and tyrosine residues through sequential activation of protein kinases. Recent studies have shown that the protein kinases involved in MAPK signal transductions might be organized into signaling complexes by scaffold proteins. These scaffold proteins are essential regulators that function by assembling the relevant molecular components in mammalian cells. In this study, we report that dual-specificity phosphatase 22 (DUSP22), a member of the protein tyrosine phosphatase family, acts as a distinct scaffold protein in c-Jun N-terminal kinase (JNK) signaling. DUSP22 increased the phosphorylation in the activation loop of JNK regardless of its phosphatase activity but had no effect on phosphorylation levels of ERK and p38 in mammalian cells. Furthermore, DUSP22 selectively associated with apoptosis signal-regulating kinase 1 (ASK1), MAPK kinase 7 (MKK7), and JNK1/2. Both JNK phosphorylation and JNK-mediated apoptosis increased in a concentration-dependent manner regardless of DUSP22 phosphatase activity at low DUSP22 concentrations, but then decreased at higher DUSP22 concentrations, which is the prominent feature of a scaffold protein. Thus, our data suggest that DUSP22 regulates cell death by acting as a scaffold protein for the ASK1-MKK7-JNK signal transduction pathway independently of its phosphatase activity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Scaffold role of a mitogen-activated protein kinase phosphatase, SKRP1, for the JNK signaling pathway.J Biol Chem. 2002 Jun 28;277(26):23919-26. doi: 10.1074/jbc.M200838200. Epub 2002 Apr 16. J Biol Chem. 2002. PMID: 11959862
-
Apoptosis signal-regulating kinase 1-mediated signaling pathway regulates hydrogen peroxide-induced apoptosis in human pulmonary vascular endothelial cells.Crit Care Med. 2003 Dec;31(12):2776-81. doi: 10.1097/01.CCM.0000098027.49562.29. Crit Care Med. 2003. PMID: 14668614
-
Cyclophilin A regulates JNK/p38-MAPK signaling through its physical interaction with ASK1.Biochem Biophys Res Commun. 2015 Aug 14;464(1):112-7. doi: 10.1016/j.bbrc.2015.06.078. Epub 2015 Jun 19. Biochem Biophys Res Commun. 2015. PMID: 26095851
-
The Dual-Specificity Phosphatase 10 (DUSP10): Its Role in Cancer, Inflammation, and Immunity.Int J Mol Sci. 2019 Apr 1;20(7):1626. doi: 10.3390/ijms20071626. Int J Mol Sci. 2019. PMID: 30939861 Free PMC article. Review.
-
Physiological roles of ASK1-mediated signal transduction in oxidative stress- and endoplasmic reticulum stress-induced apoptosis: advanced findings from ASK1 knockout mice.Antioxid Redox Signal. 2002 Jun;4(3):415-25. doi: 10.1089/15230860260196218. Antioxid Redox Signal. 2002. PMID: 12215209 Review.
Cited by
-
Dual specificity phosphatase 22 suppresses mesangial cell hyperproliferation, fibrosis, inflammation and the MAPK signaling pathway in diabetic nephropathy.Exp Ther Med. 2022 Nov 7;24(6):744. doi: 10.3892/etm.2022.11680. eCollection 2022 Dec. Exp Ther Med. 2022. PMID: 36561966 Free PMC article.
-
Phosphorylation Dynamics of JNK Signaling: Effects of Dual-Specificity Phosphatases (DUSPs) on the JNK Pathway.Int J Mol Sci. 2019 Dec 6;20(24):6157. doi: 10.3390/ijms20246157. Int J Mol Sci. 2019. PMID: 31817617 Free PMC article. Review.
-
Scaffolding of Mitogen-Activated Protein Kinase Signaling by β-Arrestins.Int J Mol Sci. 2022 Jan 17;23(2):1000. doi: 10.3390/ijms23021000. Int J Mol Sci. 2022. PMID: 35055186 Free PMC article. Review.
-
Recurrent MSC E116K mutations in ALK-negative anaplastic large cell lymphoma.Blood. 2019 Jun 27;133(26):2776-2789. doi: 10.1182/blood.2019000626. Epub 2019 May 17. Blood. 2019. PMID: 31101622 Free PMC article.
-
Modulating phosphatase DUSP22 with BML-260 ameliorates skeletal muscle wasting via Akt independent JNK-FOXO3a repression.EMBO Mol Med. 2025 Jun;17(6):1259-1288. doi: 10.1038/s44321-025-00234-2. Epub 2025 Apr 22. EMBO Mol Med. 2025. PMID: 40263624 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
