

Differential transcriptional responses to Ebola and Marburg virus infection in bat and human cells
- PMID: 27713552
- PMCID: PMC5054393
- DOI: 10.1038/srep34589
Differential transcriptional responses to Ebola and Marburg virus infection in bat and human cells
Erratum in
-
Corrigendum: Differential transcriptional responses to Ebola and Marburg virus infection in bat and human cells.Sci Rep. 2017 Jan 11;7:39421. doi: 10.1038/srep39421. Sci Rep. 2017. PMID: 28074834 Free PMC article. No abstract available.
Abstract
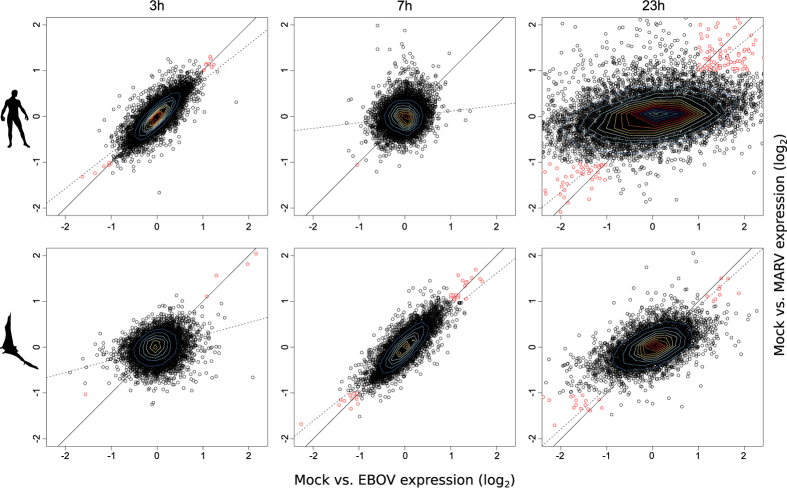
The unprecedented outbreak of Ebola in West Africa resulted in over 28,000 cases and 11,000 deaths, underlining the need for a better understanding of the biology of this highly pathogenic virus to develop specific counter strategies. Two filoviruses, the Ebola and Marburg viruses, result in a severe and often fatal infection in humans. However, bats are natural hosts and survive filovirus infections without obvious symptoms. The molecular basis of this striking difference in the response to filovirus infections is not well understood. We report a systematic overview of differentially expressed genes, activity motifs and pathways in human and bat cells infected with the Ebola and Marburg viruses, and we demonstrate that the replication of filoviruses is more rapid in human cells than in bat cells. We also found that the most strongly regulated genes upon filovirus infection are chemokine ligands and transcription factors. We observed a strong induction of the JAK/STAT pathway, of several genes encoding inhibitors of MAP kinases (DUSP genes) and of PPP1R15A, which is involved in ER stress-induced cell death. We used comparative transcriptomics to provide a data resource that can be used to identify cellular responses that might allow bats to survive filovirus infections.
Figures







References
-
- Feldmann H., Klenk H. D. & Sanchez A. Molecular biology and evolution of filoviruses. Arch. Virol. Suppl. 7, 81–100 (1993). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
