

Comparative Evaluation of Biochemical Changes in Tomato (Lycopersicon esculentum Mill.) Infected by Alternaria alternata and Its Toxic Metabolites (TeA, AOH, and AME)
- PMID: 27713751
- PMCID: PMC5031594
- DOI: 10.3389/fpls.2016.01408
Comparative Evaluation of Biochemical Changes in Tomato (Lycopersicon esculentum Mill.) Infected by Alternaria alternata and Its Toxic Metabolites (TeA, AOH, and AME)
Abstract
In the present study, we have evaluated the comparative biochemical defense response generated against Alternaria alternata and its purified toxins viz. alternariol (AOH), alternariol monomethyl ether (AME), and tenuazonic acid (TeA). The necrotic lesions developed due to treatment with toxins were almost similar as those produced by the pathogen, indicating the crucial role of these toxins in plant pathogenesis. An oxidative burst reaction characterized by the rapid and transient production of a large amount of reactive oxygen species (ROS) occurs following the pathogen infection/toxin exposure. The maximum concentration of hydrogen peroxide (H2O2) produced was reported in the pathogen infected samples (22.2-fold) at 24 h post inoculation followed by TeA (18.2-fold), AOH (15.9-fold), and AME (14.1-fold) in treated tissues. 3,3'- Diaminobenzidine staining predicted the possible sites of H2O2 accumulation while the extent of cell death was measured by Evans blue dye. The extent of lipid peroxidation and malondialdehyde (MDA) content was higher (15.8-fold) at 48 h in the sample of inoculated leaves of the pathogen when compared to control. The cellular damages were observed as increased MDA content and reduced chlorophyll. The activities of antioxidative defense enzymes increased in both the pathogen infected as well as toxin treated samples. Superoxide dismutase (SOD) activity was 5.9-fold higher at 24 h post inoculation in leaves followed by TeA (5.0-fold), AOH (4.1-fold) and AME (2.3-fold) treated leaves than control. Catalase (CAT) activity was found to be increased upto 48 h post inoculation and maximum in the pathogen challenged samples followed by other toxins. The native PAGE results showed the variations in the intensities of isozyme (SOD and CAT) bands in the pathogen infected and toxin treated samples. Ascorbate peroxidase (APx) and glutathione reductase (GR) activities followed the similar trend to scavenge the excess H2O2. The reduction in CAT activities after 48 h post inoculation demonstrate that the biochemical defense programming shown by the host against the pathogen is not well efficient resulting in the compatible host-pathogen interaction. The elicitor (toxins) induced biochemical changes depends on the potential toxic effects (extent of ROS accumulation, amount of H2O2 produced). Thus, a fine tuning occurs for the defense related antioxidative enzymes against detoxification of key ROS molecules and effectively regulated in tomato plant against the pathogen infected/toxin treated oxidative stress. The study well demonstrates the acute pathological effects of A. alternata in tomato over its phytotoxic metabolites.
Keywords: Alternaria alternata; Polymerase chain reaction (PCR); pathogen; phytotoxic metabolites; toxins.
Figures
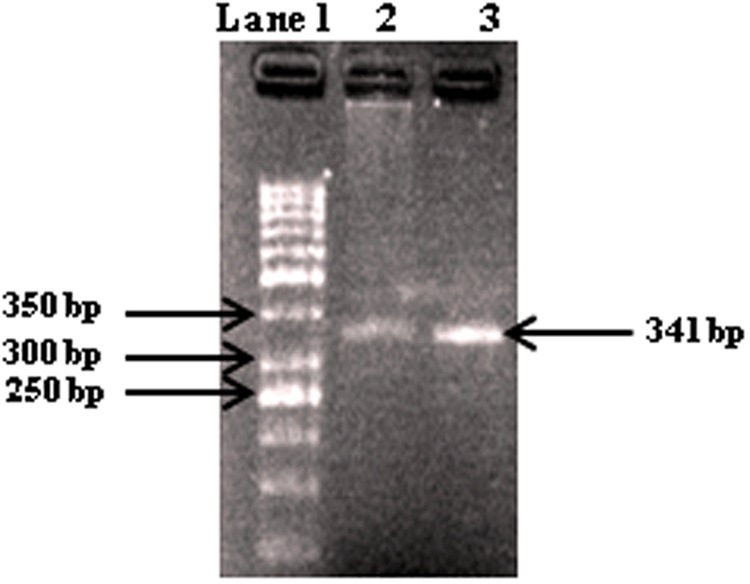


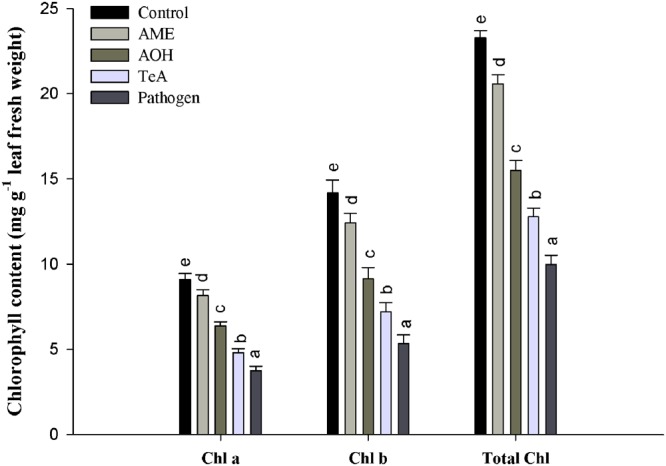

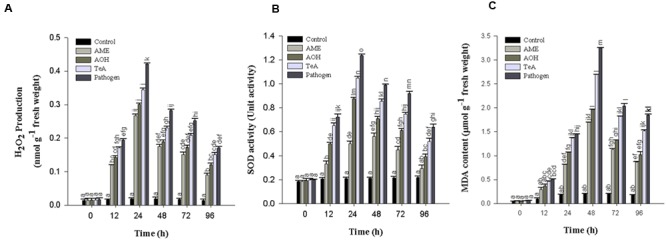
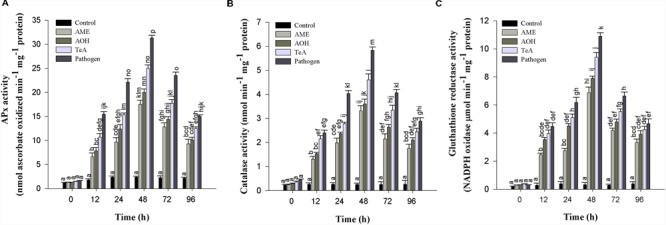
References
-
- Abd-El Kareem F., El-Mougy N. S., El-Gamal N. G., Fatouh Y. O. (2006). Use of chitin and chitisan against tomato root rot disease under greenhouse conditions. Res. J. Agric. Biol. Sci. 2 147–152.
-
- Ackermann Y., Curtui V., Dietrich R., Gross M., Latif H., Märtlbauer E., et al. (2011). Widespread occurrence of low levels of alternariol in apple and tomato products, as determinants by comparative immunochemical assessment using monoclonal and polyclonal antibodies. J. Agric. Food Chem. 59 6360–6368. 10.1021/jf201516f - DOI - PubMed
-
- Aebi H. (1984). “Catalase,” in Methods in Enzymology Vol. 105 ed. Packer L. (Orlando, FL: Academic Press; ) 121–126. - PubMed
-
- Agrawal G. K., Jwa N. S., Rakwal R. (2002). A pathogen-induced novel rice (Oryza sativa L.) gene encodes a putative protein homologous to type II glutathione-S-transferases. Plant Sci. 163 1153–1160. 10.1016/S0168-9452(02)00331-X - DOI
-
- Agrios G. N. (2005). Plant Pathology. Burlington, MA: Elsevier Academic Press.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous