

Oligodendrocyte Development in the Absence of Their Target Axons In Vivo
- PMID: 27716830
- PMCID: PMC5055324
- DOI: 10.1371/journal.pone.0164432
Oligodendrocyte Development in the Absence of Their Target Axons In Vivo
Abstract
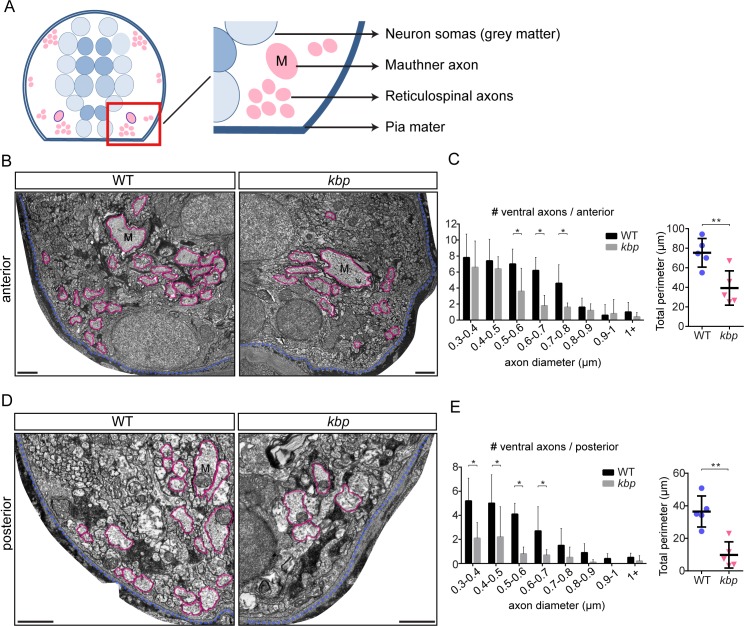
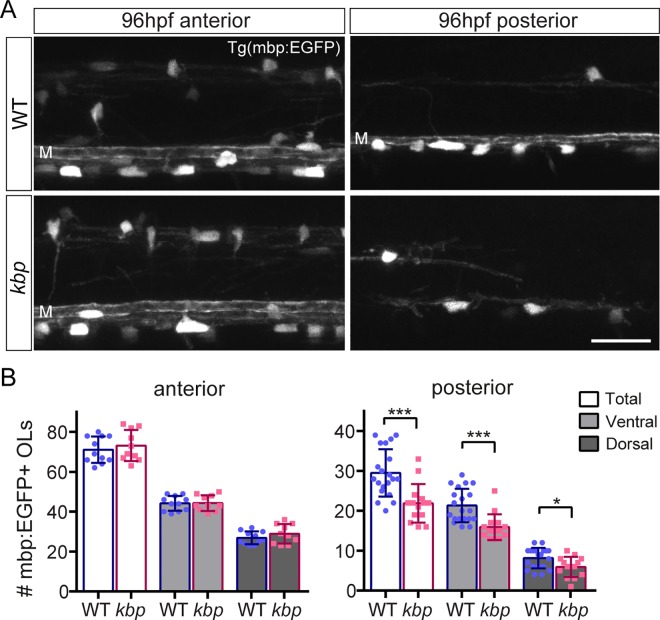
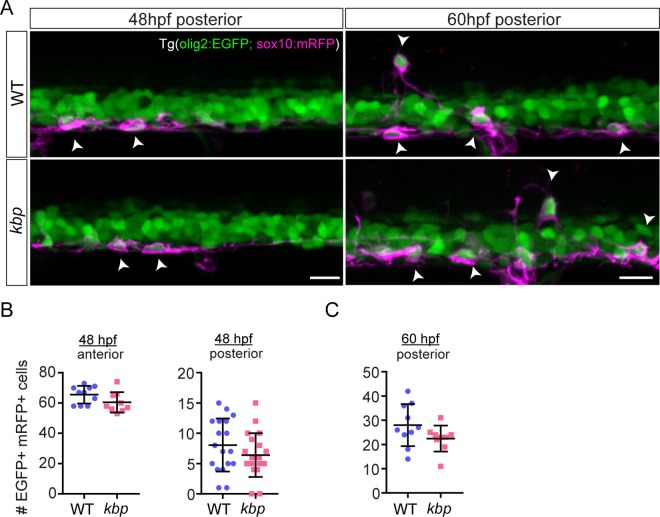
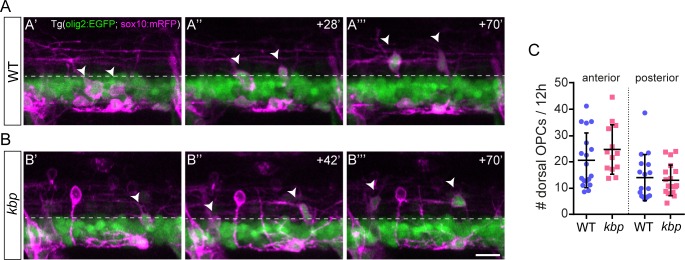
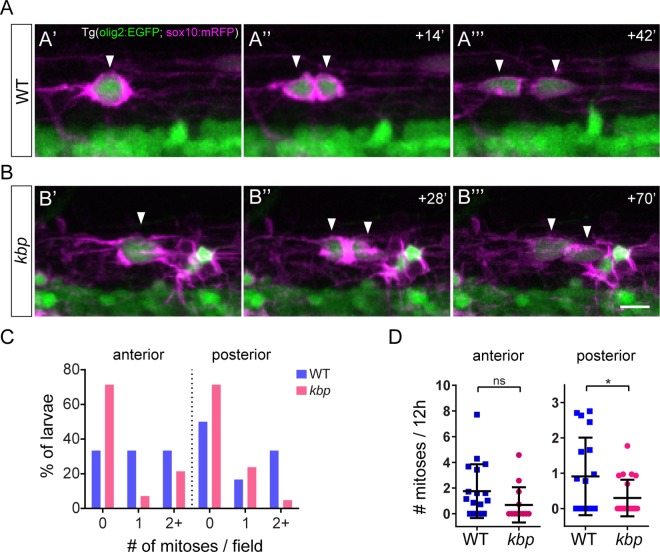
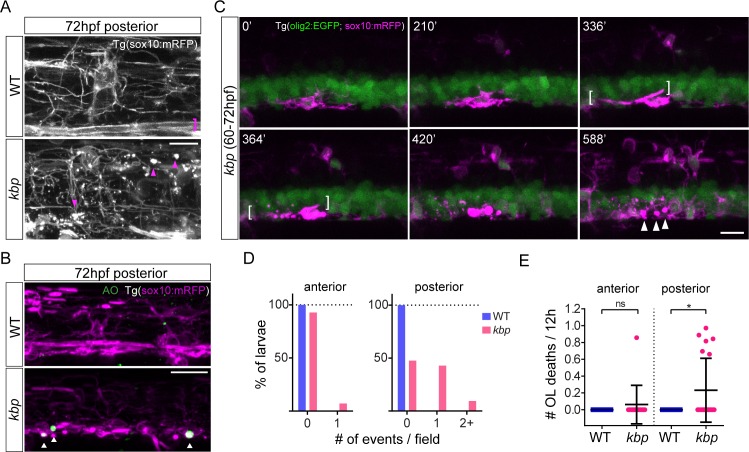
Oligodendrocytes form myelin around axons of the central nervous system, enabling saltatory conduction. Recent work has established that axons can regulate certain aspects of oligodendrocyte development and myelination, yet remarkably oligodendrocytes in culture retain the ability to differentiate in the absence of axons and elaborate myelin sheaths around synthetic axon-like substrates. It remains unclear the extent to which the life-course of oligodendrocytes requires the presence of, or signals derived from axons in vivo. In particular, it is unclear whether the specific axons fated for myelination regulate the oligodendrocyte population in a living organism, and if so, which precise steps of oligodendrocyte-cell lineage progression are regulated by target axons. Here, we use live-imaging of zebrafish larvae carrying transgenic reporters that label oligodendrocyte-lineage cells to investigate which aspects of oligodendrocyte development, from specification to differentiation, are affected when we manipulate the target axonal environment. To drastically reduce the number of axons targeted for myelination, we use a previously identified kinesin-binding protein (kbp) mutant, in which the first myelinated axons in the spinal cord, reticulospinal axons, do not fully grow in length, creating a region in the posterior spinal cord where most initial targets for myelination are absent. We find that a 73% reduction of reticulospinal axon surface in the posterior spinal cord of kbp mutants results in a 27% reduction in the number of oligodendrocytes. By time-lapse analysis of transgenic OPC reporters, we find that the reduction in oligodendrocyte number is explained by a reduction in OPC proliferation and survival. Interestingly, OPC specification and migration are unaltered in the near absence of normal axonal targets. Finally, we find that timely differentiation of OPCs into oligodendrocytes does not depend at all on the presence of target axons. Together, our data illustrate the power of zebrafish for studying the entire life-course of the oligodendrocyte lineage in vivo in an altered axonal environment.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
