

Death Receptor 5 Networks Require Membrane Cholesterol for Proper Structure and Function
- PMID: 27720987
- PMCID: PMC5394796
- DOI: 10.1016/j.jmb.2016.10.001
Death Receptor 5 Networks Require Membrane Cholesterol for Proper Structure and Function
Abstract
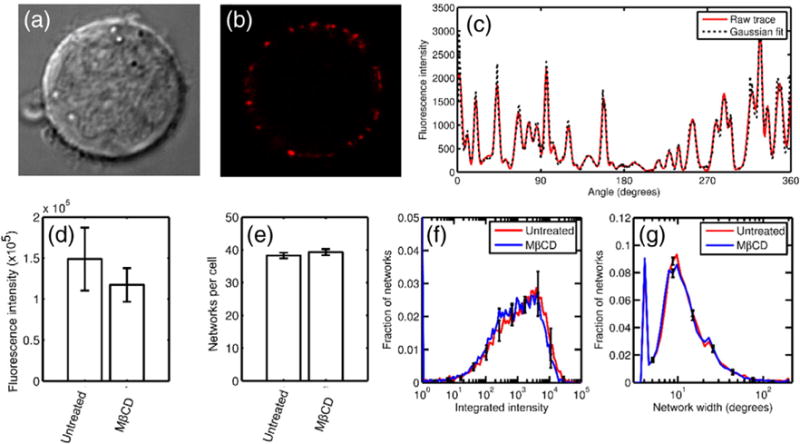
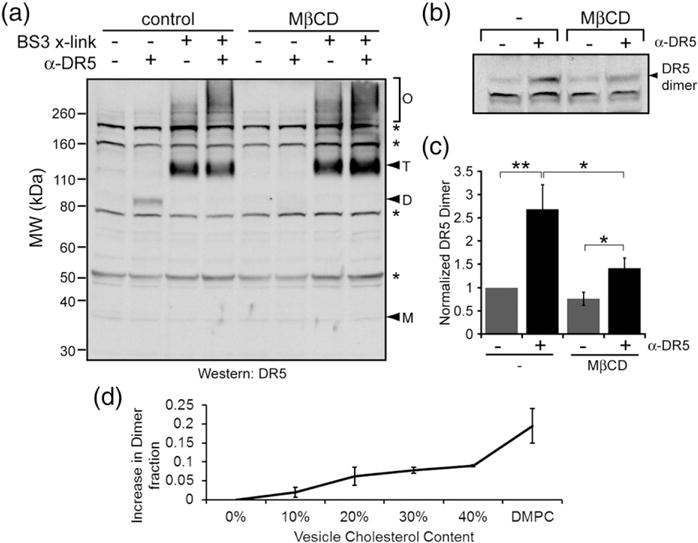
Death receptor 5 (DR5) is an apoptosis-inducing member of the tumor necrosis factor receptor superfamily, whose activity has been linked to membrane cholesterol content. Upon ligand binding, DR5 forms large clusters within the plasma membrane that have often been assumed to be manifestations of receptor co-localization in cholesterol-rich membrane domains. However, we have recently shown that DR5 clusters are more than just randomly aggregated receptors. Instead, these are highly structured networks held together by receptor dimers. These dimers are stabilized by specific transmembrane helix-helix interactions, including a disulfide bond in the long isoform of the receptor. The complex relationships among DR5 network formation, transmembrane helix dimerization, membrane cholesterol, and receptor activity has not been established. It is unknown whether the membrane itself plays an active role in driving DR5 transmembrane helix interactions or in the formation of the networks. We show that cholesterol depletion in cells does not inhibit the formation of DR5 networks. However, the networks that form in cholesterol-depleted cells fail to induce caspase cleavage. These results suggest a potential structural difference between active and inactive networks. As evidence, we show that cholesterol is necessary for the covalent dimerization of DR5 transmembrane domains. Molecular simulations and experiments in synthetic vesicles on the DR5 transmembrane dimer suggest that dimerization is facilitated by increased helicity in a thicker bilayer.
Keywords: cholesterol-rich membrane domains; disulfide bond; ligand/receptor clustering; replica exchange molecular dynamics; transmembrane domain.
Copyright © 2016. Published by Elsevier Ltd.
Figures



Similar articles
-
Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) induces death receptor 5 networks that are highly organized.J Biol Chem. 2012 Jun 15;287(25):21265-78. doi: 10.1074/jbc.M111.306480. Epub 2012 Apr 10. J Biol Chem. 2012. PMID: 22496450 Free PMC article.
-
The conformation of the extracellular binding domain of Death Receptor 5 in the presence and absence of the activating ligand TRAIL: a molecular dynamics study.Proteins. 2008 Feb 1;70(2):333-43. doi: 10.1002/prot.21541. Proteins. 2008. PMID: 17671978
-
Higher-Order Clustering of the Transmembrane Anchor of DR5 Drives Signaling.Cell. 2019 Mar 7;176(6):1477-1489.e14. doi: 10.1016/j.cell.2019.02.001. Epub 2019 Feb 28. Cell. 2019. PMID: 30827683 Free PMC article.
-
Two death-inducing human TRAIL receptors to target in cancer: similar or distinct regulation and function?Biochem Pharmacol. 2014 Oct 15;91(4):447-56. doi: 10.1016/j.bcp.2014.08.010. Epub 2014 Aug 20. Biochem Pharmacol. 2014. PMID: 25150214 Review.
-
Functional marriage in plasma membrane: Critical cholesterol level-optimal protein activity.Br J Pharmacol. 2020 Jun;177(11):2456-2465. doi: 10.1111/bph.15027. Epub 2020 Mar 24. Br J Pharmacol. 2020. PMID: 32060896 Free PMC article. Review.
Cited by
-
Death Receptor 5 Activation Is Energetically Coupled to Opening of the Transmembrane Domain Dimer.Biophys J. 2017 Jul 25;113(2):381-392. doi: 10.1016/j.bpj.2017.05.038. Biophys J. 2017. PMID: 28746849 Free PMC article.
-
Conformational states of TNFR1 as a molecular switch for receptor function.Protein Sci. 2020 Jun;29(6):1401-1415. doi: 10.1002/pro.3829. Epub 2020 Jan 31. Protein Sci. 2020. PMID: 31960514 Free PMC article.
-
Cholesterol restricts lymphotoxin β receptor-triggered NF-κB signaling.Cell Commun Signal. 2019 Dec 26;17(1):171. doi: 10.1186/s12964-019-0460-1. Cell Commun Signal. 2019. PMID: 31878945 Free PMC article.
-
Fast-diffusing p75NTR monomers support apoptosis and growth cone collapse by neurotrophin ligands.Proc Natl Acad Sci U S A. 2019 Oct 22;116(43):21563-21572. doi: 10.1073/pnas.1902790116. Epub 2019 Sep 12. Proc Natl Acad Sci U S A. 2019. PMID: 31515449 Free PMC article.
-
Noncompetitive Allosteric Antagonism of Death Receptor 5 by a Synthetic Affibody Ligand.Biochemistry. 2020 Oct 13;59(40):3856-3868. doi: 10.1021/acs.biochem.0c00529. Epub 2020 Sep 30. Biochemistry. 2020. PMID: 32941010 Free PMC article.
References
-
- Wiley SR, Schooley K, Smolak PJ, Din WS, Huang CP, Nicholl JK, et al. Identification and characterization of a new member of the TNF family that induces apoptosis. Immunity. 1995;3:673–682. - PubMed
-
- Pan G, Ni J, Wei YF, Yu G, Gentz R, Dixit VM. An antagonist decoy receptor and a death domain-containing receptor for TRAIL. Science. 1997;277:815–818. http://dx.doi.org/10.1126/science.277.5327.815. - DOI - PubMed
-
- Thorburn A. Death receptor-induced cell killing. Cell Signal. 2004;16:139–144. - PubMed
-
- Schneider P, Tschopp J. Apoptosis induced by death receptors. Pharm Acta Helv. 2000;74:281–286. - PubMed
-
- Bodmer JL, Schneider P, Tschopp J. The molecular architecture of the TNF superfamily. Trends Biochem Sci. 2002;27:19–26. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
