

Evolutionarily conserved dual lysine motif determines the non-chaperone function of secreted Hsp90alpha in tumour progression
- PMID: 27721406
- PMCID: PMC5386837
- DOI: 10.1038/onc.2016.375
Evolutionarily conserved dual lysine motif determines the non-chaperone function of secreted Hsp90alpha in tumour progression
Erratum in
-
Correction: Evolutionarily conserved dual lysine motif determines the non-chaperone function of secreted Hsp90alpha in tumour progression.Oncogene. 2024 May;43(18):1397-1398. doi: 10.1038/s41388-024-03017-0. Oncogene. 2024. PMID: 38575761 Free PMC article. No abstract available.
Abstract
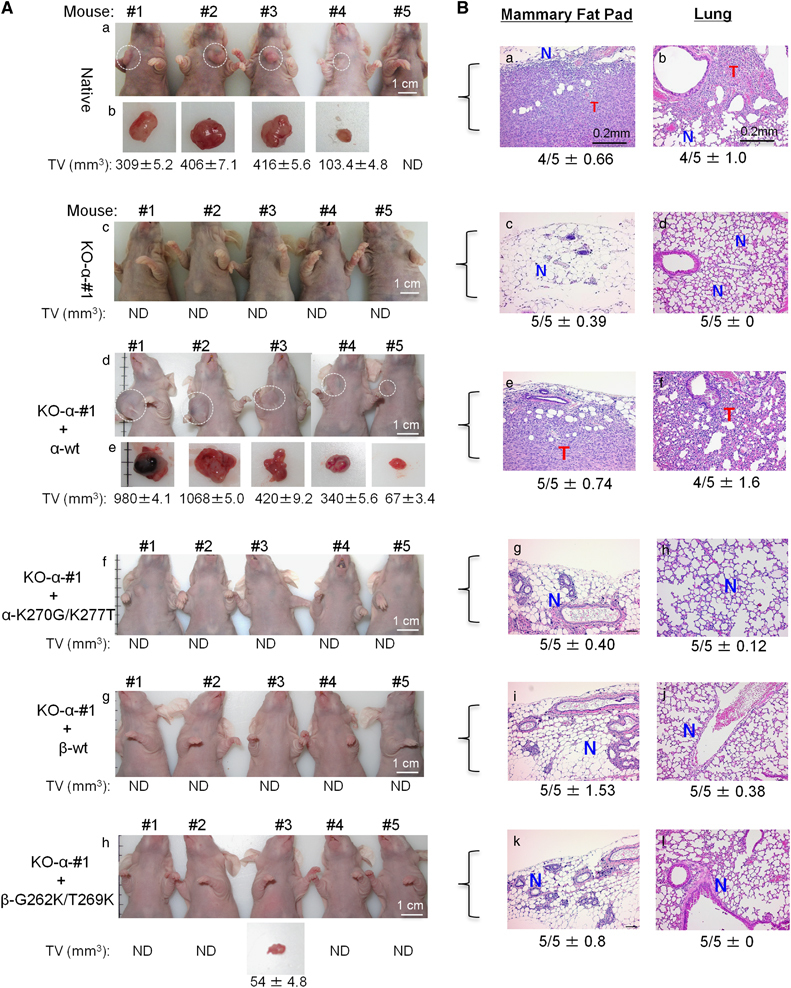
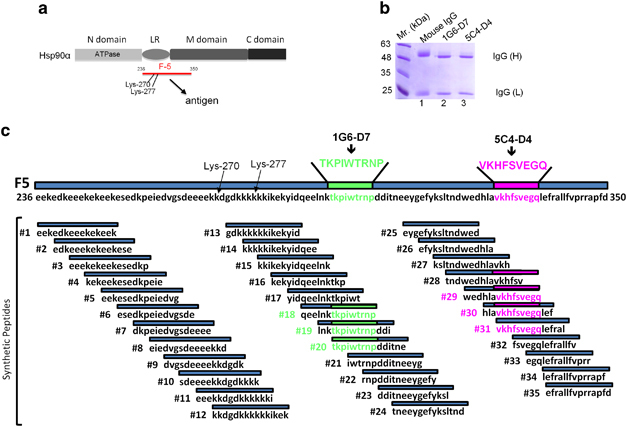
Both intracellular and extracellular heat shock protein-90 (Hsp90) family proteins (α and β) have been shown to support tumour progression. The tumour-supporting activity of the intracellular Hsp90 is attributed to their N-terminal ATPase-driven chaperone function. What molecular entity determines the extracellular function of secreted Hsp90 and the distinction between Hsp90α and Hsp90β was unclear. Here we demonstrate that CRISPR/Case9 knocking out Hsp90α nullifies tumour cells' ability to migrate, invade and metastasize without affecting the cell survival and growth. Knocking out Hsp90β leads to tumour cell death. Extracellular supplementation with recombinant Hsp90α, but not Hsp90β, protein recovers tumourigenicity of the Hsp90α-knockout cells. Sequential mutagenesis identifies two evolutionarily conserved lysine residues, lys-270 and lys-277, in the Hsp90α subfamily that determine the extracellular Hsp90α function. Hsp90β subfamily lacks the dual lysine motif and the extracellular function. Substitutions of gly-262 and thr-269 in Hsp90β with lysines convert Hsp90β to a Hsp90α-like protein. Newly constructed monoclonal antibody, 1G6-D7, against the dual lysine region of secreted Hsp90α inhibits both de novo tumour formation and expansion of already formed tumours in mice. This study suggests an alternative therapeutic approach to target Hsp90 in cancer, that is, the tumour-secreted Hsp90α, instead of the intracellular Hsp90α and Hsp90β.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Drysdale MJ, Brough PA, Massey A, Jensen MR, Schoepfer J . Targeting Hsp90 for the treatment of cancer. Curr Opin Drug Discov Devel 2006; 9: 483–495. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
