

Development of a stretch-induced neurotrauma model for medium-throughput screening in vitro: identification of rifampicin as a neuroprotectant
- PMID: 27723079
- PMCID: PMC5758395
- DOI: 10.1111/bph.13642
Development of a stretch-induced neurotrauma model for medium-throughput screening in vitro: identification of rifampicin as a neuroprotectant
Abstract
Background and purpose: We hypothesized that an in vitro, stretch-based model of neural injury may be useful to identify compounds that decrease the cellular damage in neurotrauma.
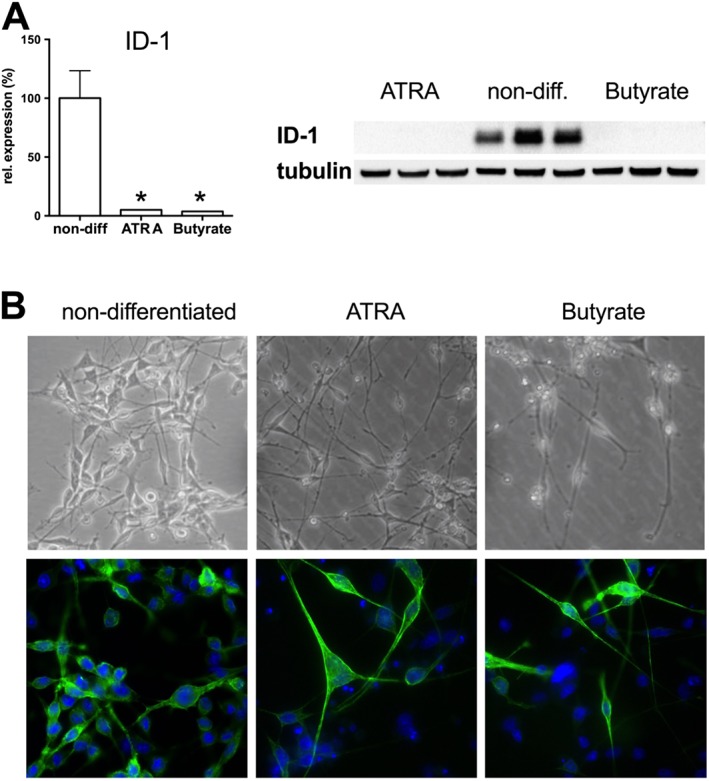
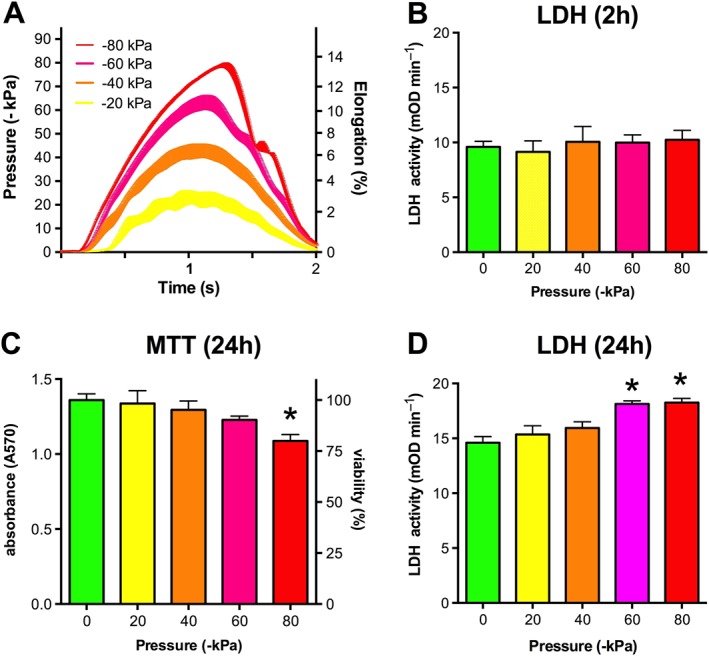
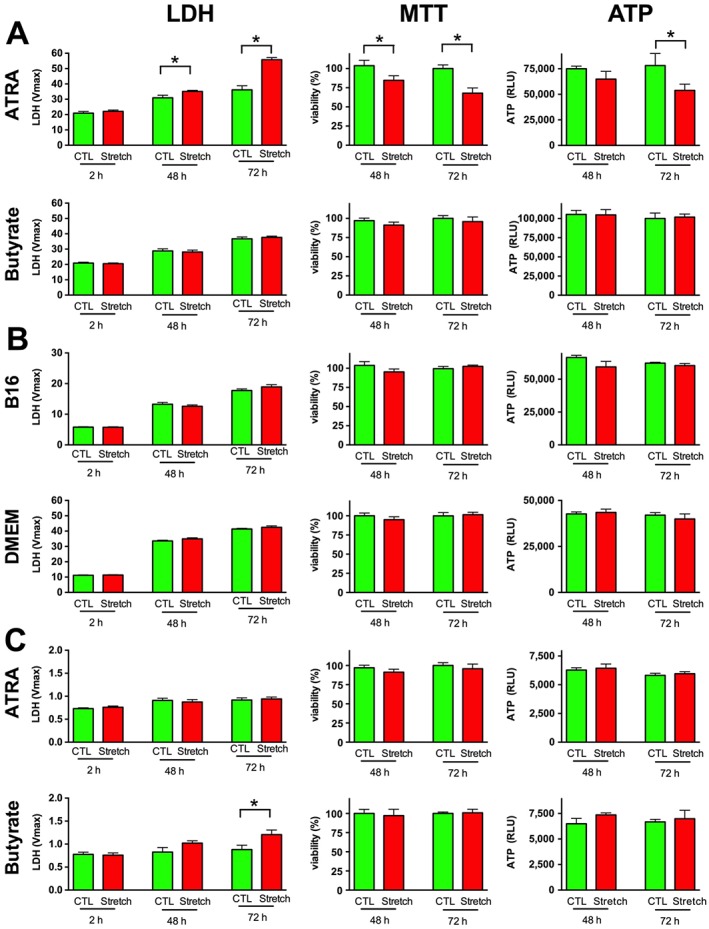
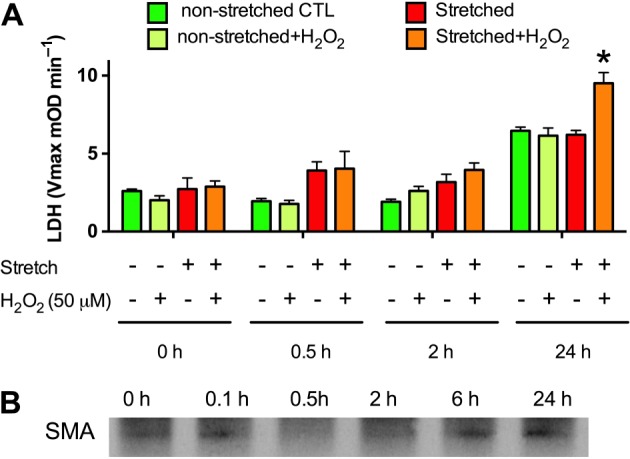
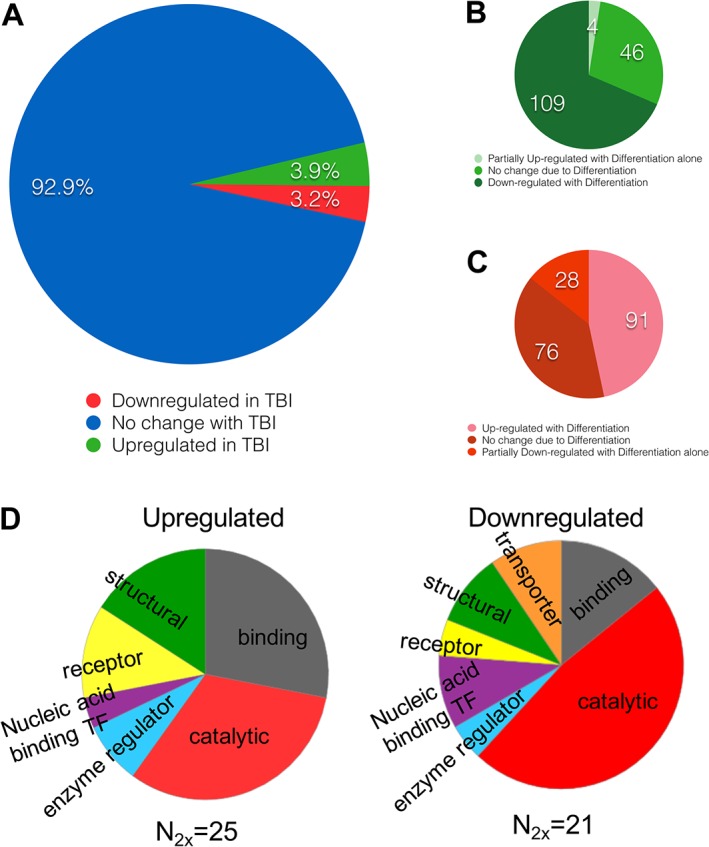
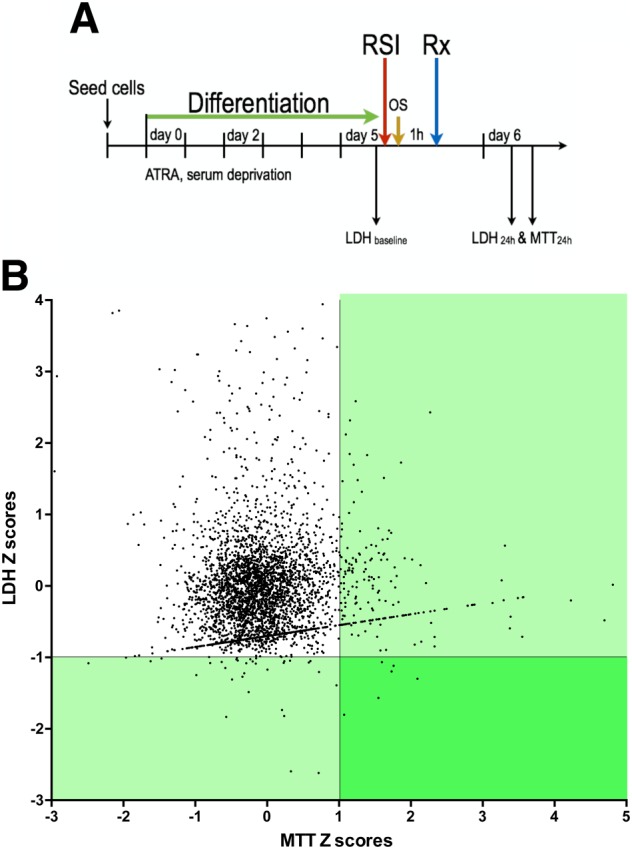
Experimental approach: We screened three neural cell lines (B35, RN33B and SH-SY5Y) subjected to two differentiation methods and selected all-trans-retinoic acid-differentiated B35 rat neuroblastoma cells subjected to rapid stretch injury, coupled with a subthreshold concentration of H2 O2 , for the screen. The model induced marked alterations in gene expression and proteomic signature of the cells and culminated in delayed cell death (LDH release) and mitochondrial dysfunction [reduced 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) conversion]. Follow-up studies utilized human stem cell-derived neurons subjected to rapid stretch injury.
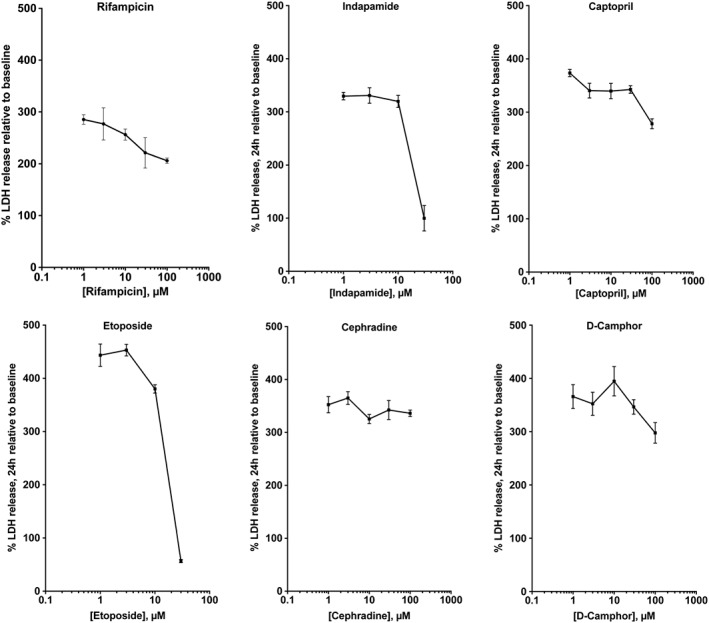
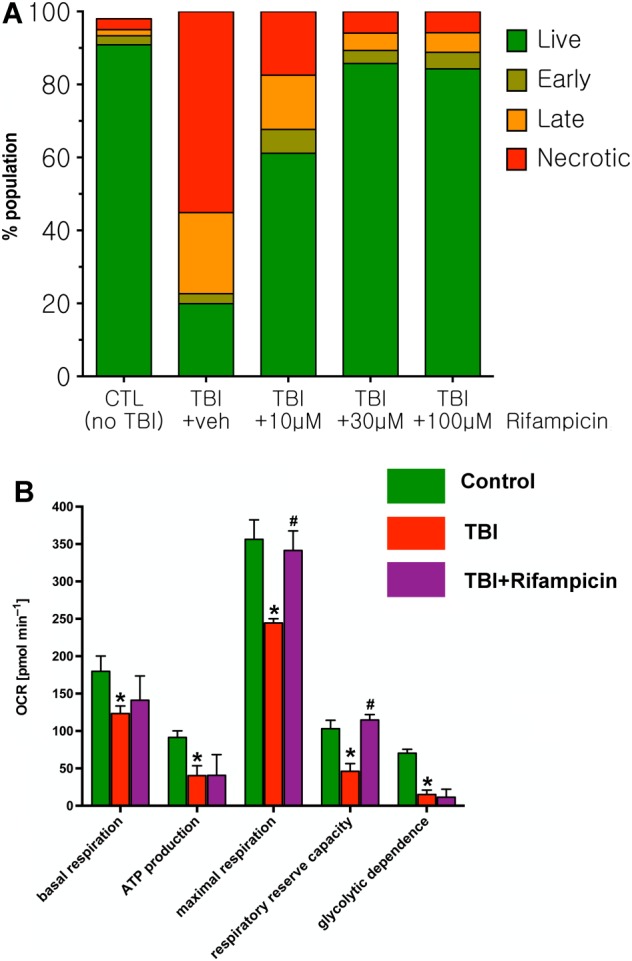
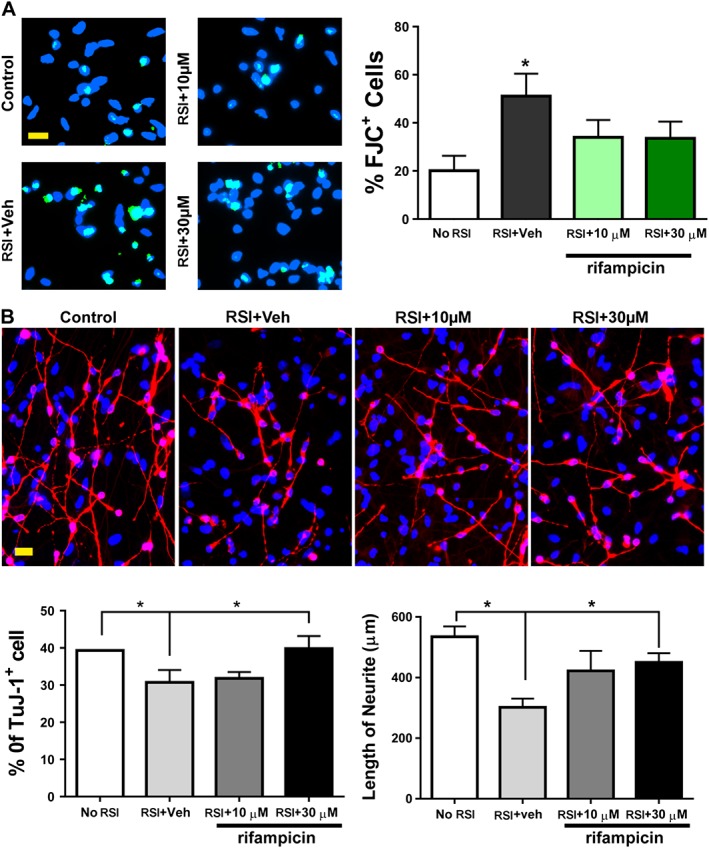
Key results: From screening of a composite library of 3500 drugs, five drugs (when applied in a post-treatment regimen relative to stretch injury) improved both LDH and MTT responses. The effects of rifampicin were investigated in further detail. Rifampicin reduced cell necrosis and apoptosis and improved cellular bioenergetics. In a second model (stretch injury in human stem cell-derived neurons), rifampicin pretreatment attenuated LDH release, protected against the loss of neurite length and maintained neuron-specific class III β-tubulin immunoreactivity.
Conclusions and implications: We conclude that the current model is suitable for medium-throughput screening to identify compounds with neuroprotective potential. Rifampicin, when applied either in pre- or post-treatment, improves the viability of neurons subjected to stretch injury and protects against neurite loss. Rifampicin may be a candidate for repurposing for the therapy of traumatic brain injury.
Linked articles: This article is part of a themed section on Inventing New Therapies Without Reinventing the Wheel: The Power of Drug Repurposing. To view the other articles in this section visit http://onlinelibrary.wiley.com/doi/10.1111/bph.v175.2/issuetoc.
© 2016 The British Pharmacological Society.
Figures

References
-
- Bell JD, Park E, Ai J, Baker AJ (2009). PICK1‐mediated GluR2 endocytosis contributes to cellular injury after neuronal trauma. Cell Death Differ 16: 1665–1680. - PubMed
-
- Biagini G, Zoli M, Torri C, Boschi S, Vantaggiato G, Ballestri M et al. (1997). Protective effects of delapril, indapamide and their combination chronically administered to stroke‐prone spontaneously hypertensive rats fed a high‐sodium diet. Clin Sci (Lond) 93: 401–411. - PubMed
-
- Bramlett HM, Dietrich WD (2007). Progressive damage after brain and spinal cord injury: pathomechanisms and treatment strategies. Prog Brain Res 161: 125–124. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
