

An excitatory cortical feedback loop gates retinal wave transmission in rodent thalamus
- PMID: 27725086
- PMCID: PMC5059135
- DOI: 10.7554/eLife.18816
An excitatory cortical feedback loop gates retinal wave transmission in rodent thalamus
Abstract
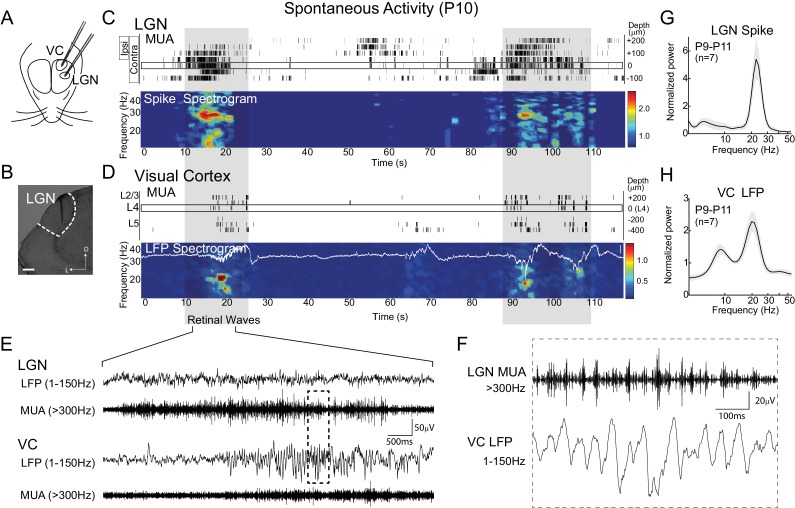
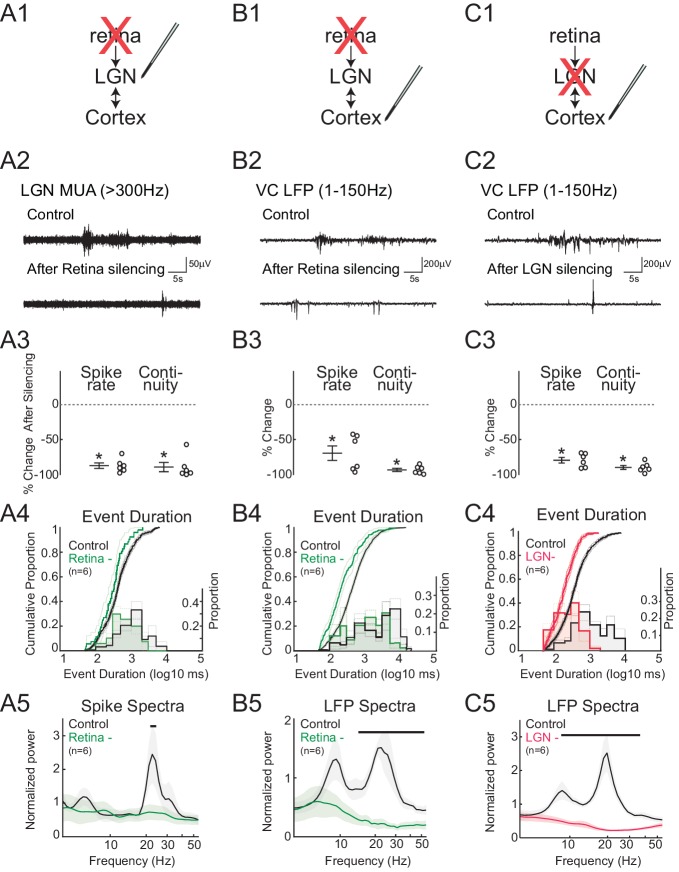
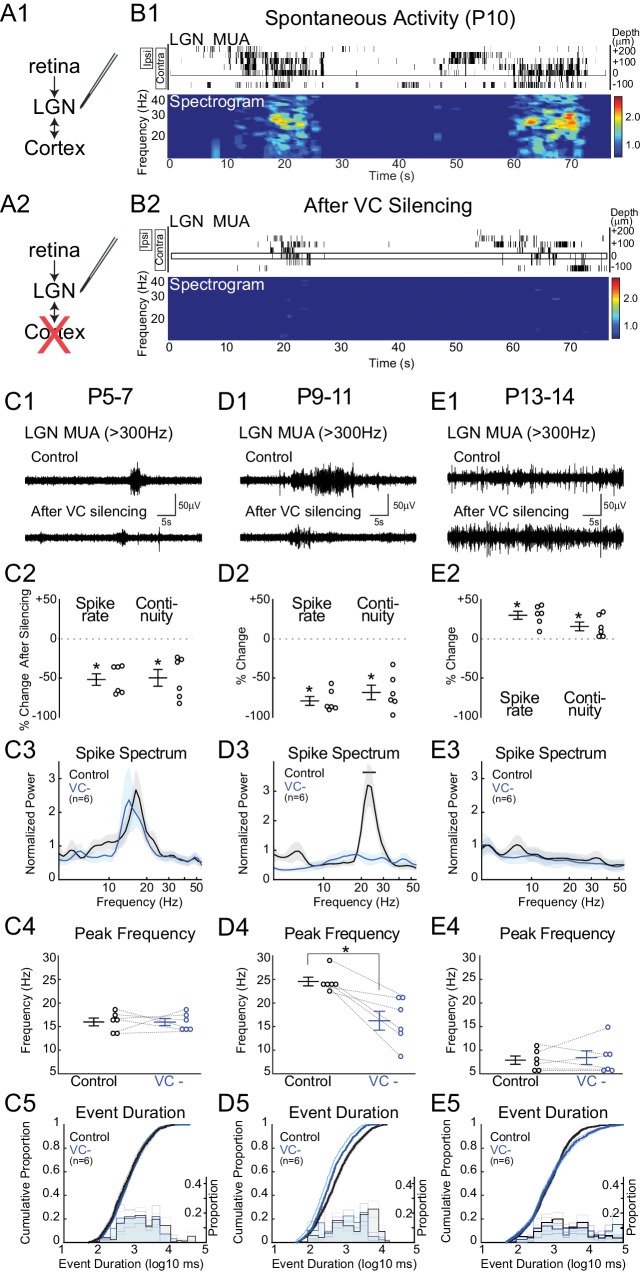
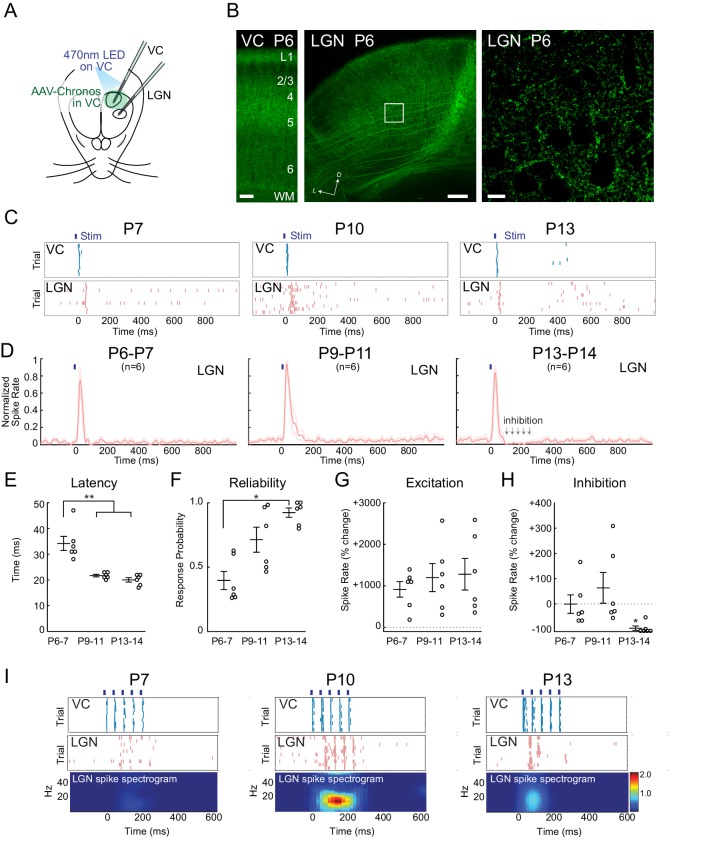
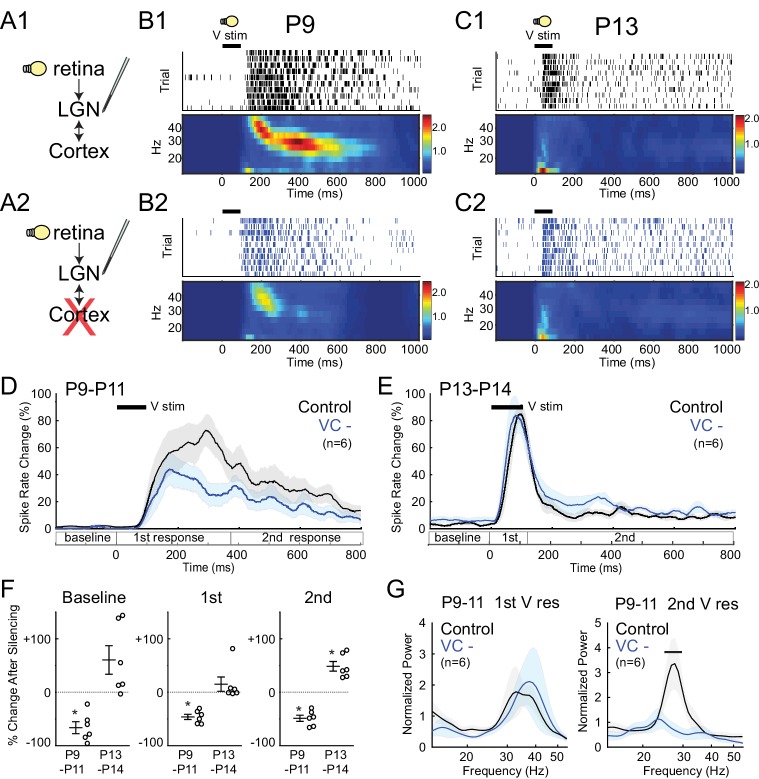
Spontaneous retinal waves are critical for the development of receptive fields in visual thalamus (LGN) and cortex (VC). Despite a detailed understanding of the circuit specializations in retina that generate waves, whether central circuit specializations also exist to control their propagation through visual pathways of the brain is unknown. Here we identify a developmentally transient, corticothalamic amplification of retinal drive to thalamus as a mechanism for retinal wave transmission in the infant rat brain. During the period of retinal waves, corticothalamic connections excite LGN, rather than driving feedforward inhibition as observed in the adult. This creates an excitatory feedback loop that gates retinal wave transmission through the LGN. This cortical multiplication of retinal wave input ends just prior to eye-opening, as cortex begins to inhibit LGN. Our results show that the early retino-thalamo-cortical circuit uses developmentally specialized feedback amplification to ensure powerful, high-fidelity transmission of retinal activity despite immature connectivity.
Keywords: EEG; activity depedent development; neuroscience; oscillations; plasticity; rat; retinal waves.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
Comment in
-
Catching a wave.Elife. 2016 Oct 11;5:e21236. doi: 10.7554/eLife.21236. Elife. 2016. PMID: 27725090 Free PMC article.
Similar articles
-
Retinal and Nonretinal Contributions to Extraclassical Surround Suppression in the Lateral Geniculate Nucleus.J Neurosci. 2017 Jan 4;37(1):226-235. doi: 10.1523/JNEUROSCI.1577-16.2016. J Neurosci. 2017. PMID: 28053044 Free PMC article.
-
Feed-forward synchronization: propagation of temporal patterns along the retinothalamocortical pathway.Philos Trans R Soc Lond B Biol Sci. 2002 Dec 29;357(1428):1869-76. doi: 10.1098/rstb.2002.1172. Philos Trans R Soc Lond B Biol Sci. 2002. PMID: 12626020 Free PMC article. Review.
-
Retinal waves trigger spindle bursts in the neonatal rat visual cortex.J Neurosci. 2006 Jun 21;26(25):6728-36. doi: 10.1523/JNEUROSCI.0752-06.2006. J Neurosci. 2006. PMID: 16793880 Free PMC article.
-
Increasing Spontaneous Retinal Activity before Eye Opening Accelerates the Development of Geniculate Receptive Fields.J Neurosci. 2015 Oct 28;35(43):14612-23. doi: 10.1523/JNEUROSCI.1365-15.2015. J Neurosci. 2015. PMID: 26511250 Free PMC article.
-
The influence of the corticothalamic projection on responses in thalamus and cortex.Philos Trans R Soc Lond B Biol Sci. 2002 Dec 29;357(1428):1823-34. doi: 10.1098/rstb.2002.1159. Philos Trans R Soc Lond B Biol Sci. 2002. PMID: 12626015 Free PMC article. Review.
Cited by
-
Polynomial, piecewise-Linear, Step (PLS): A Simple, Scalable, and Efficient Framework for Modeling Neurons.Front Neuroinform. 2021 May 6;15:642933. doi: 10.3389/fninf.2021.642933. eCollection 2021. Front Neuroinform. 2021. PMID: 34025382 Free PMC article.
-
Synaptic and circuit mechanisms prevent detrimentally precise correlation in the developing mammalian visual system.Elife. 2023 May 22;12:e84333. doi: 10.7554/eLife.84333. Elife. 2023. PMID: 37211984 Free PMC article.
-
Spontaneous activity in developing thalamic and cortical sensory networks.Neuron. 2021 Aug 18;109(16):2519-2534. doi: 10.1016/j.neuron.2021.06.026. Epub 2021 Jul 21. Neuron. 2021. PMID: 34293296 Free PMC article. Review.
-
The Integrative Function of Silent Synapses on Subplate Neurons in Cortical Development and Dysfunction.Front Neuroanat. 2019 Apr 16;13:41. doi: 10.3389/fnana.2019.00041. eCollection 2019. Front Neuroanat. 2019. PMID: 31040772 Free PMC article. Review.
-
Thalamocortical circuits for the formation of hierarchical pathways in the mammalian visual cortex.Front Neural Circuits. 2023 Apr 17;17:1155195. doi: 10.3389/fncir.2023.1155195. eCollection 2023. Front Neural Circuits. 2023. PMID: 37139079 Free PMC article. Review.
References
-
- André M, Lamblin M-D, d’Allest AM, Curzi-Dascalova L, Moussalli-Salefranque F, Nguyen The Tich S, Vecchierini-Blineau M-F, Wallois F, Walls-Esquivel E, Plouin P. Electroencephalography in premature and full-term infants. Developmental features and glossary. Neurophysiologie Clinique/Clinical Neurophysiology. 2010;40:59–124. doi: 10.1016/j.neucli.2010.02.002. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
