

Mitochondrial Superoxide Signaling Contributes to Norepinephrine-Mediated T-Lymphocyte Cytokine Profiles
- PMID: 27727316
- PMCID: PMC5058488
- DOI: 10.1371/journal.pone.0164609
Mitochondrial Superoxide Signaling Contributes to Norepinephrine-Mediated T-Lymphocyte Cytokine Profiles
Abstract
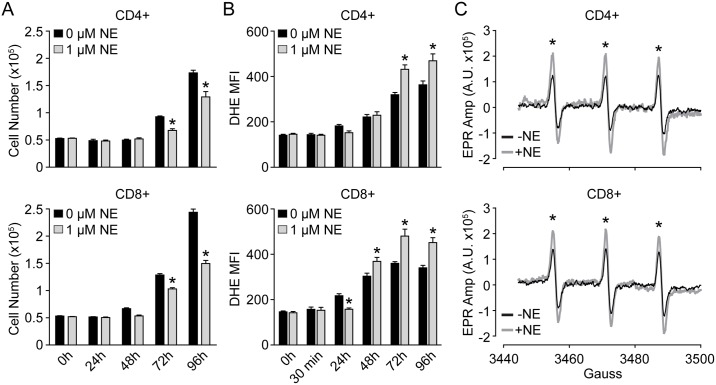
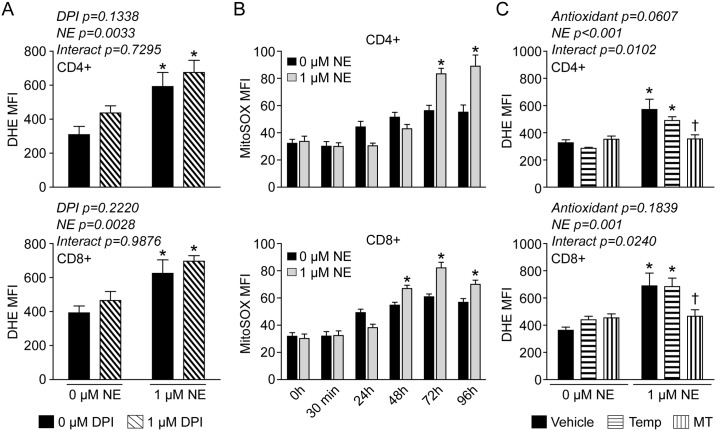
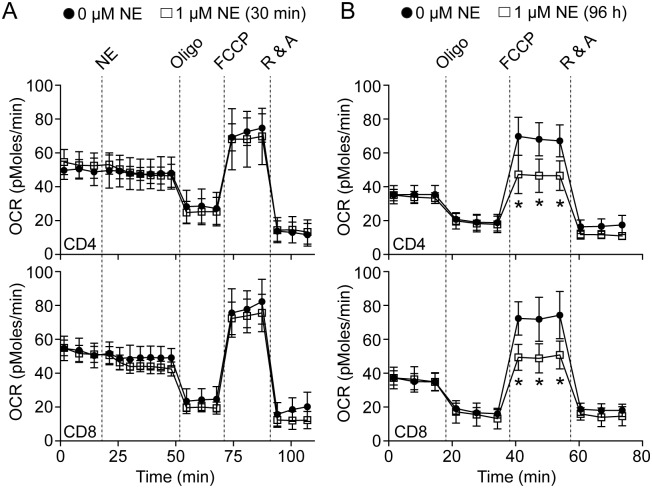
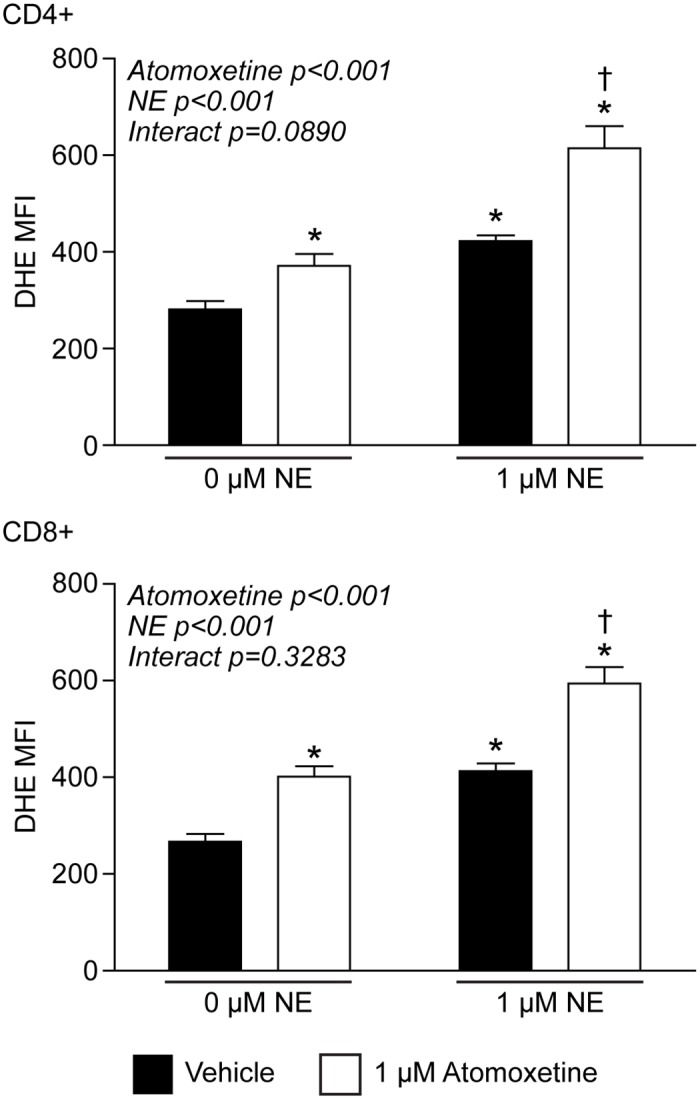
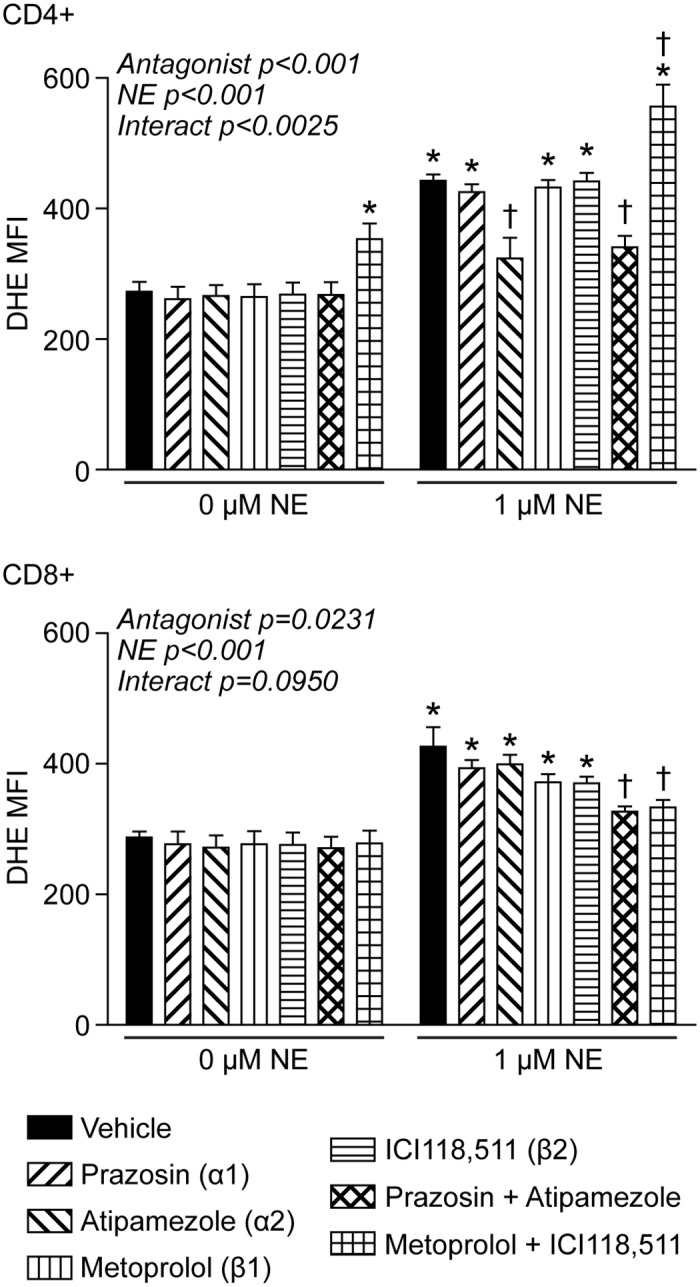
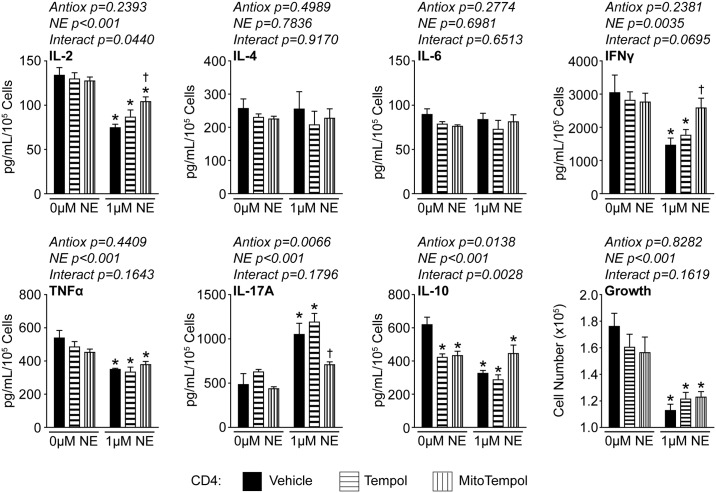
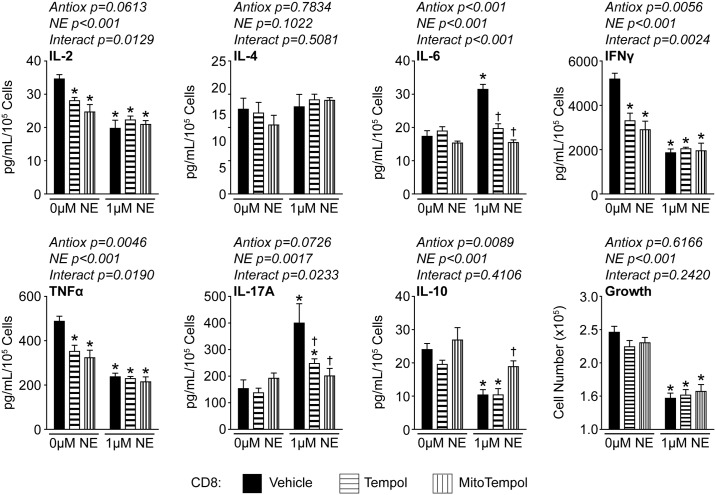
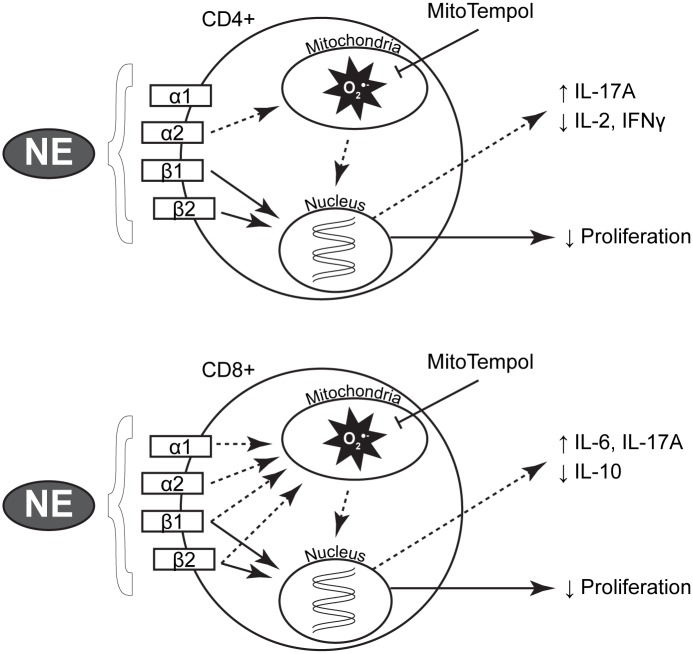
Norepinephrine (NE) produces multifaceted regulatory patterns in T-lymphocytes. Recently, we have shown that NE utilizes redox signaling as evidenced by increased superoxide (O2●-) causally linked to the observed changes in these cells; however, the source of this reactive oxygen species (ROS) remains elusive. Herein, we hypothesized that the source of increased O2●- in NE-stimulated T-lymphocytes is due to disruption of mitochondrial bioenergetics. To address this hypothesis, we utilized purified mouse splenic CD4+ and CD8+ T-lymphocytes stimulated with NE and assessed O2●- levels, mitochondrial metabolism, cellular proliferation, and cytokine profiles. We demonstrate that the increase in O2●- levels in response to NE is time-dependent and occurs at later points of T-lymphocyte activation. Moreover, the source of O2●- was indeed the mitochondria as evidenced by enhanced MitoSOX Red oxidation as well as abrogation of this signal by the addition of the mitochondrial-targeted O2●--scavenging antioxidant MitoTempol. NE-stimulated T-lymphocytes also demonstrated decreased mitochondrial respiratory capacity, which suggests disruption of mitochondrial metabolism and the potential source of increased mitochondrial O2●-. The effects of NE in regards to redox signaling appear to be adrenergic receptor-dependent as specific receptor antagonists could reverse the increase in O2●-; however, differential receptors regulating these processes were observed in CD4+ versus CD8+ T-lymphocytes. Finally, mitochondrial O2●- was shown to be mechanistic to the NE-mediated T-lymphocyte phenotype as supplementation of MitoTempol could reverse specific changes in cytokine expression observed with NE treatment. Overall, these studies indicate that mitochondrial metabolism and O2●--mediated redox signaling play a regulatory role in the T-lymphocyte response to NE.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
