

Retinol and ascorbate drive erasure of epigenetic memory and enhance reprogramming to naïve pluripotency by complementary mechanisms
- PMID: 27729528
- PMCID: PMC5086989
- DOI: 10.1073/pnas.1608679113
Retinol and ascorbate drive erasure of epigenetic memory and enhance reprogramming to naïve pluripotency by complementary mechanisms
Abstract
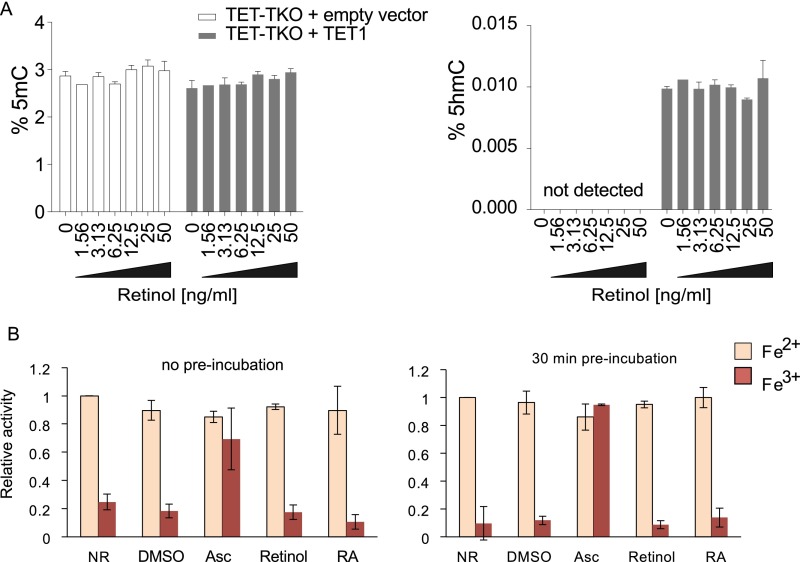
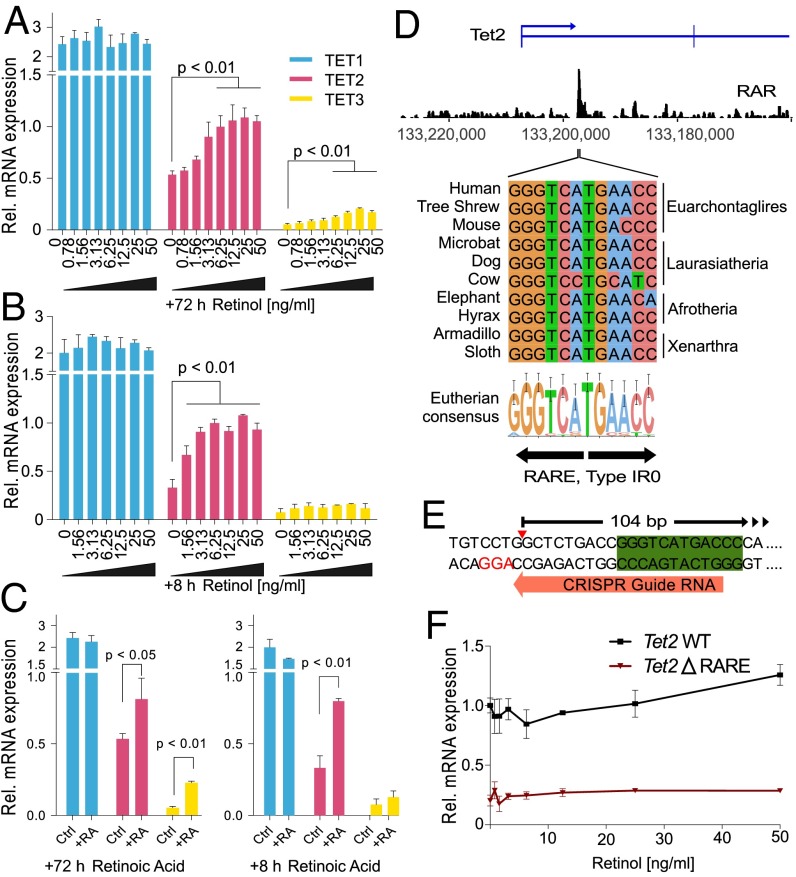
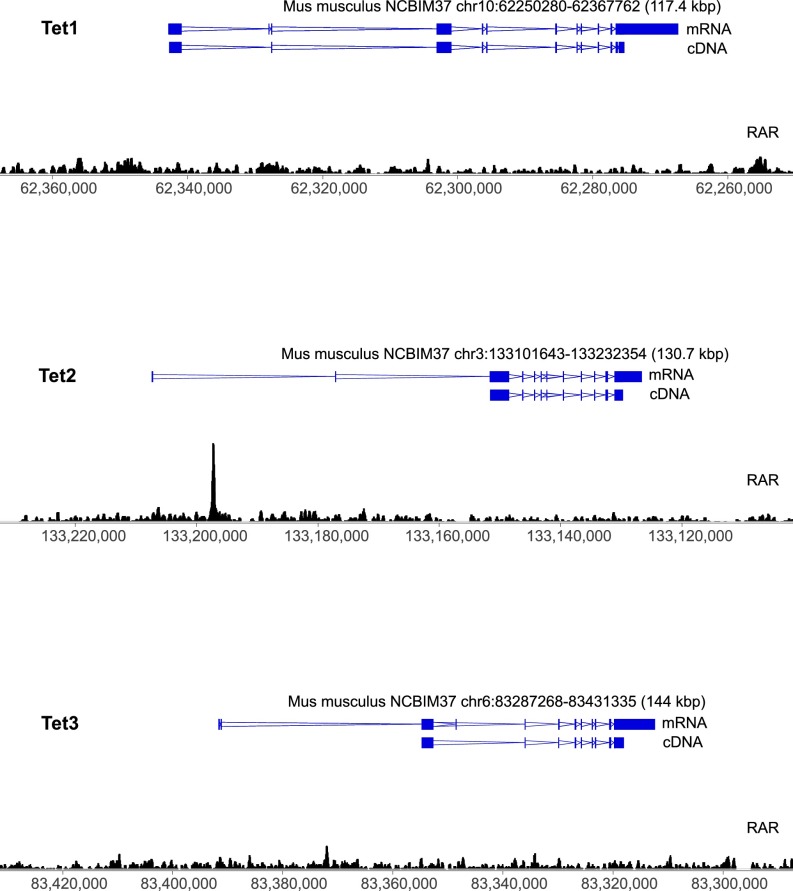
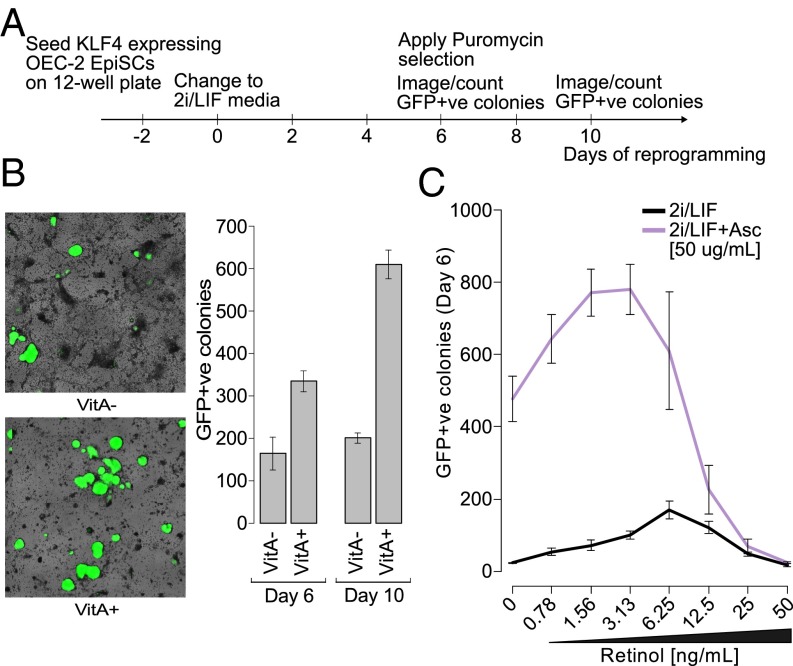
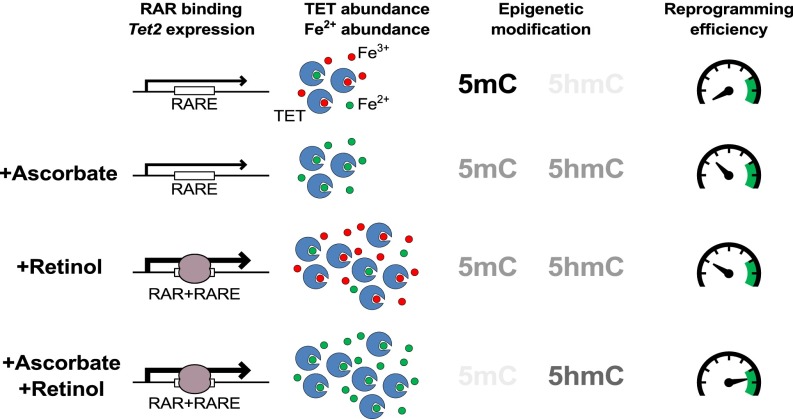
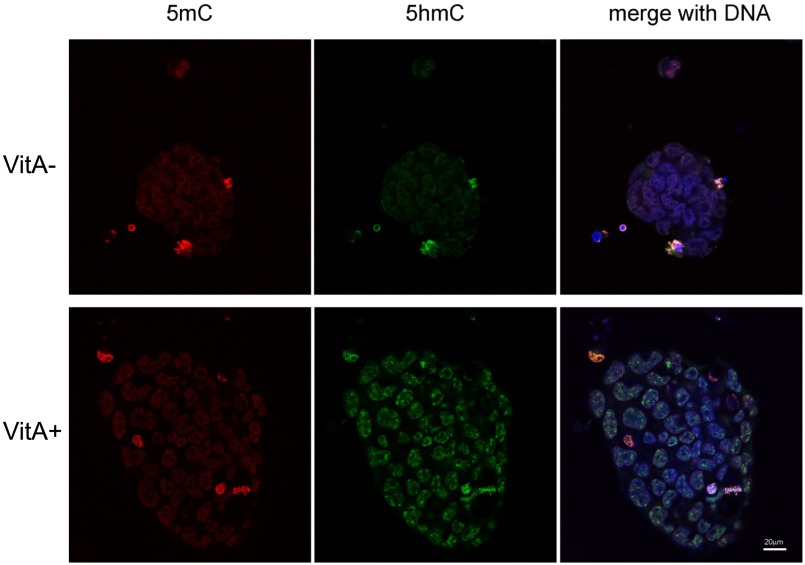
Epigenetic memory, in particular DNA methylation, is established during development in differentiating cells and must be erased to create naïve (induced) pluripotent stem cells. The ten-eleven translocation (TET) enzymes can catalyze the oxidation of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC) and further oxidized derivatives, thereby actively removing this memory. Nevertheless, the mechanism by which the TET enzymes are regulated, and the extent to which they can be manipulated, are poorly understood. Here we report that retinoic acid (RA) or retinol (vitamin A) and ascorbate (vitamin C) act as modulators of TET levels and activity. RA or retinol enhances 5hmC production in naïve embryonic stem cells by activation of TET2 and TET3 transcription, whereas ascorbate potentiates TET activity and 5hmC production through enhanced Fe2+ recycling, and not as a cofactor as reported previously. We find that both ascorbate and RA or retinol promote the derivation of induced pluripotent stem cells synergistically and enhance the erasure of epigenetic memory. This mechanistic insight has significance for the development of cell treatments for regenenerative medicine, and enhances our understanding of how intrinsic and extrinsic signals shape the epigenome.
Keywords: DNA methylation; TET; epigenetic memory; naive pluripotency; vitamin A/C.
Conflict of interest statement
W.R. and S.B. serve as consultants for Cambridge Epigenetix Ltd.
Figures
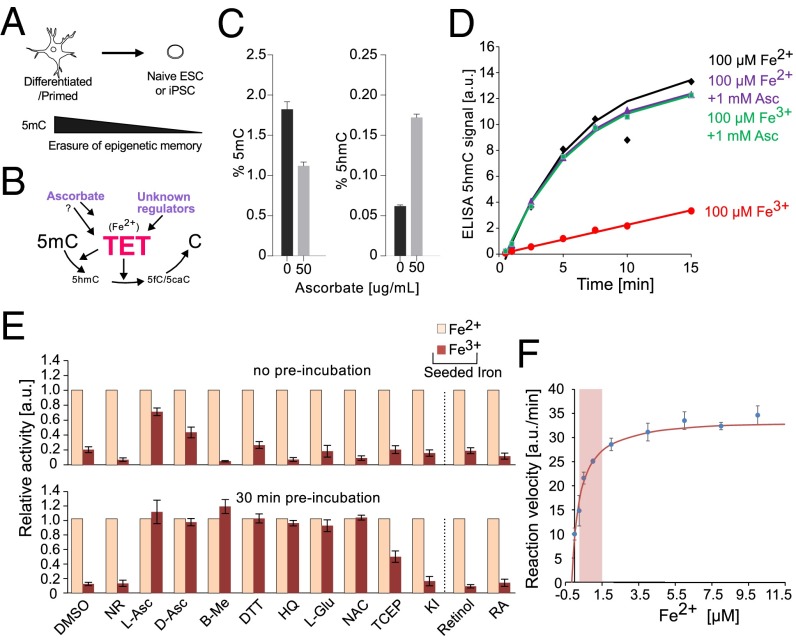
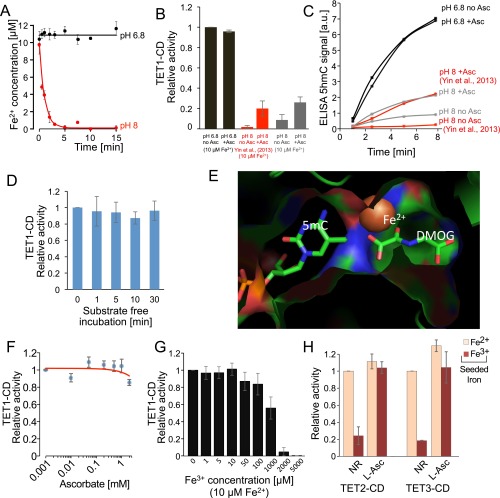
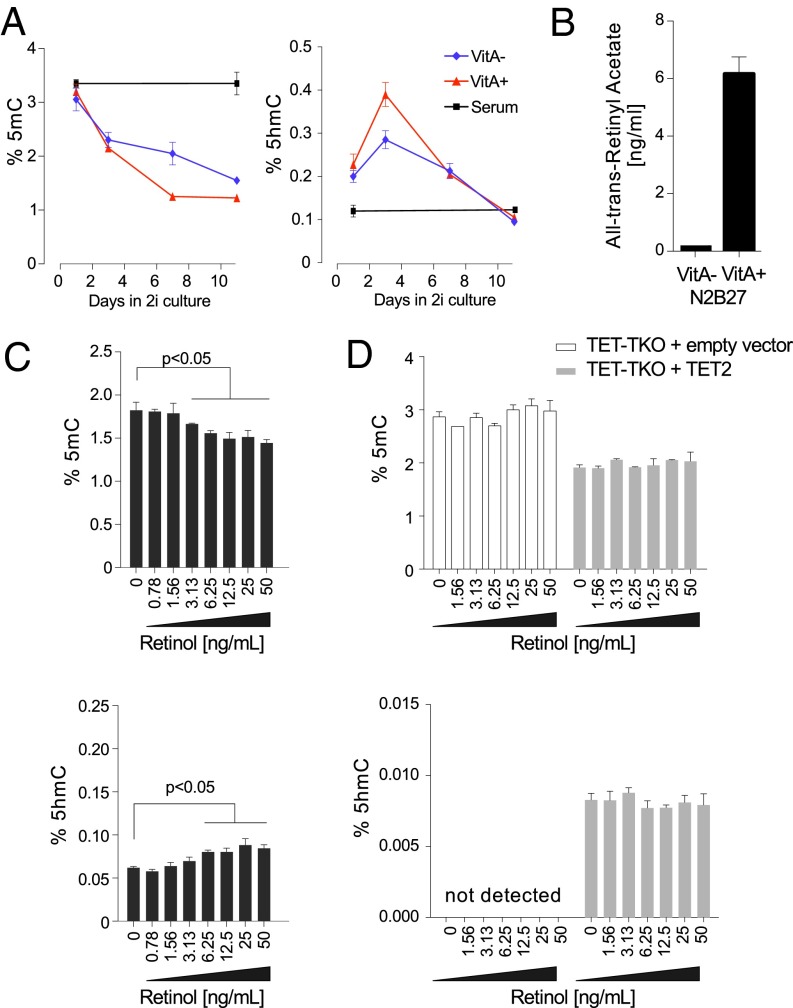
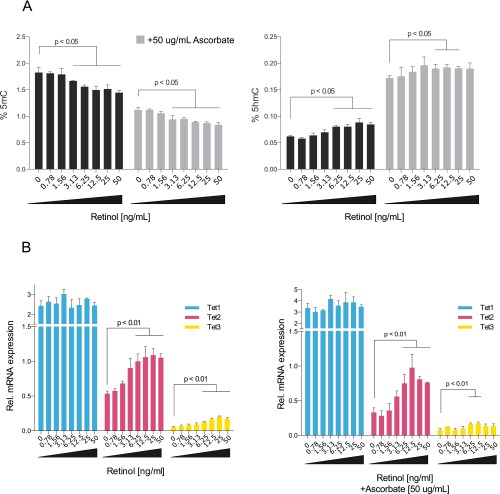
References
-
- Habibi E, et al. Whole-genome bisulfite sequencing of two distinct interconvertible DNA methylomes of mouse embryonic stem cells. Cell Stem Cell. 2013;13(3):360–369. - PubMed
-
- Yamaji M, et al. PRDM14 ensures naive pluripotency through dual regulation of signaling and epigenetic pathways in mouse embryonic stem cells. Cell Stem Cell. 2013;12(3):368–382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
