

Mechanism and disease association of E2-conjugating enzymes: lessons from UBE2T and UBE2L3
- PMID: 27729585
- PMCID: PMC5095918
- DOI: 10.1042/BCJ20160028
Mechanism and disease association of E2-conjugating enzymes: lessons from UBE2T and UBE2L3
Abstract
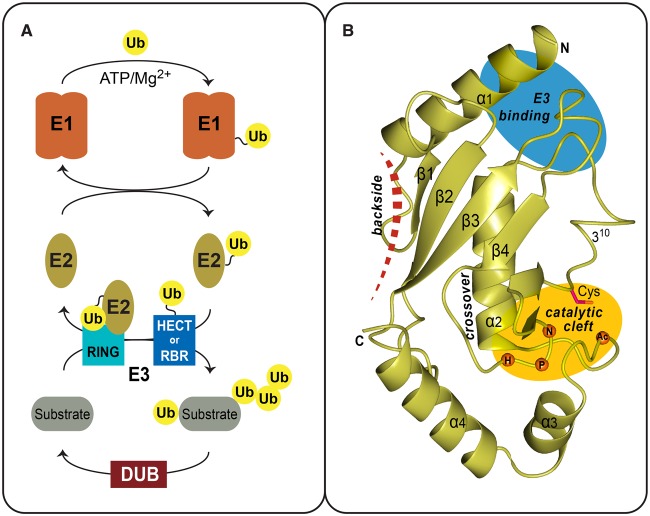
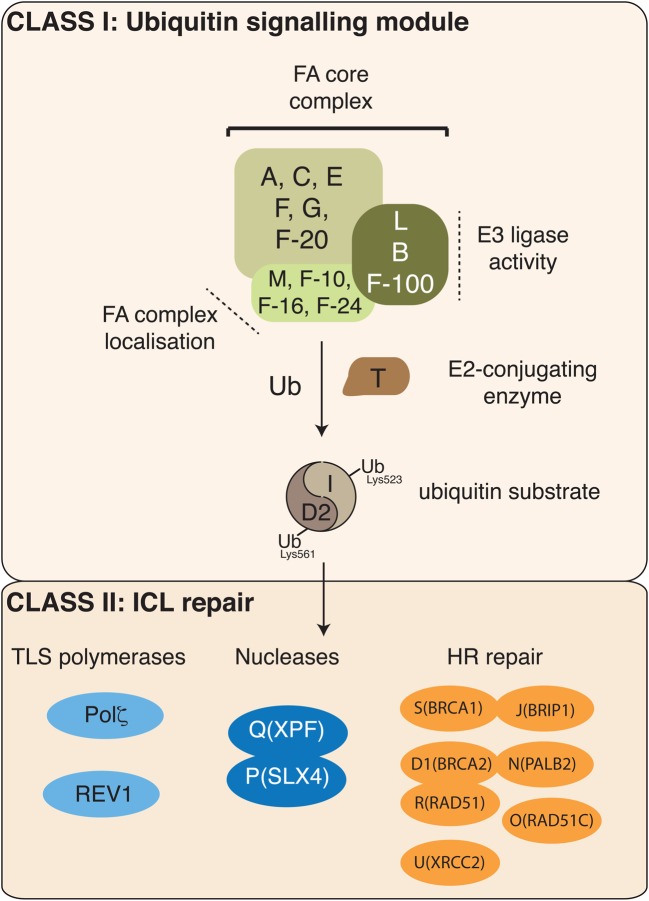
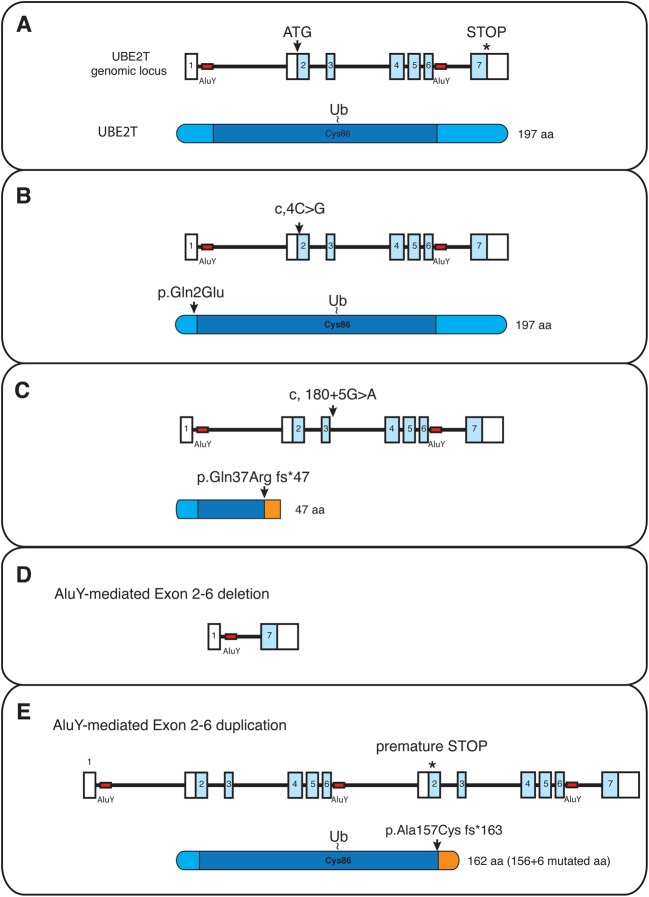
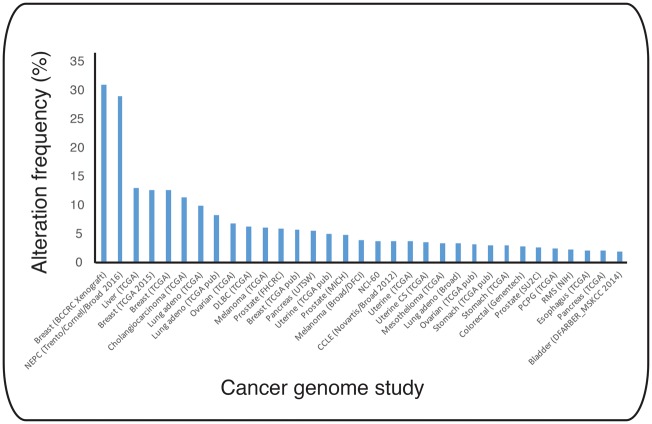
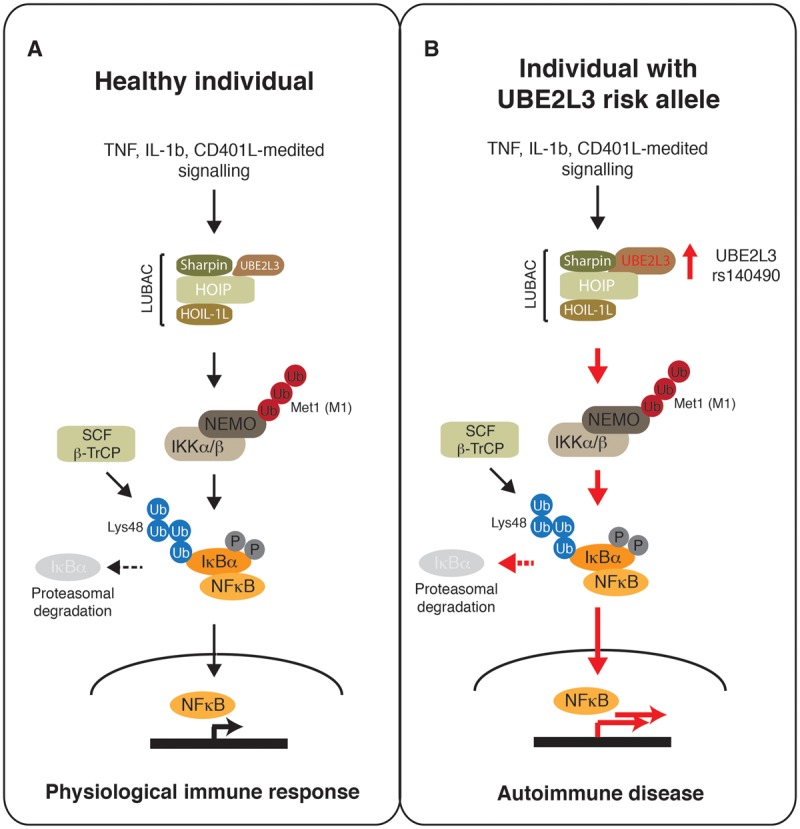
Ubiquitin signalling is a fundamental eukaryotic regulatory system, controlling diverse cellular functions. A cascade of E1, E2, and E3 enzymes is required for assembly of distinct signals, whereas an array of deubiquitinases and ubiquitin-binding modules edit, remove, and translate the signals. In the centre of this cascade sits the E2-conjugating enzyme, relaying activated ubiquitin from the E1 activating enzyme to the substrate, usually via an E3 ubiquitin ligase. Many disease states are associated with dysfunction of ubiquitin signalling, with the E3s being a particular focus. However, recent evidence demonstrates that mutations or impairment of the E2s can lead to severe disease states, including chromosome instability syndromes, cancer predisposition, and immunological disorders. Given their relevance to diseases, E2s may represent an important class of therapeutic targets. In the present study, we review the current understanding of the mechanism of this important family of enzymes, and the role of selected E2s in disease.
Keywords: E2-conjugating enzyme; Fanconi anaemia; UBE2L3; UBE2T; autoimmune disease; ubiquitin signalling.
© 2016 The Author(s).
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous