

Robust Label-free, Quantitative Profiling of Circulating Plasma Microparticle (MP) Associated Proteins
- PMID: 27738094
- PMCID: PMC5141277
- DOI: 10.1074/mcp.M116.060491
Robust Label-free, Quantitative Profiling of Circulating Plasma Microparticle (MP) Associated Proteins
Abstract
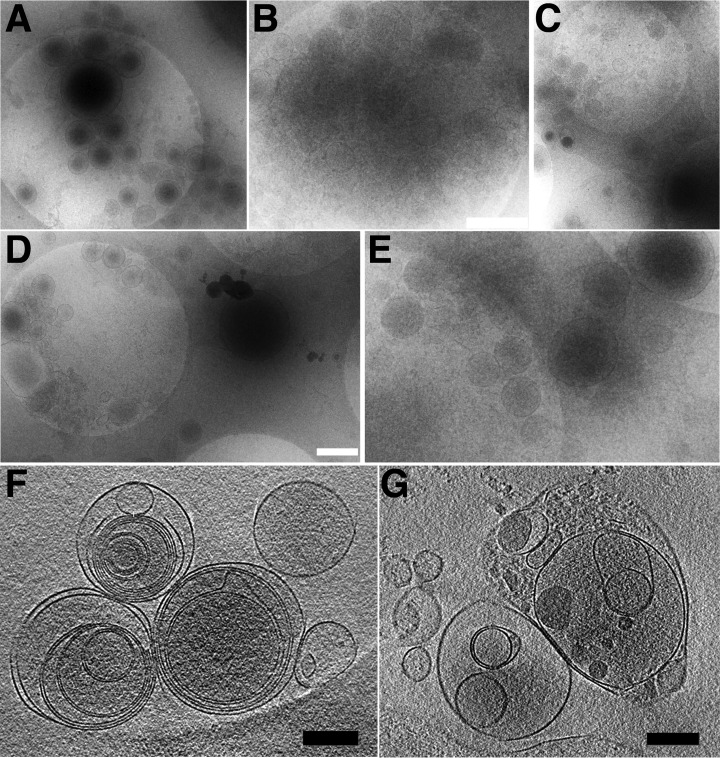
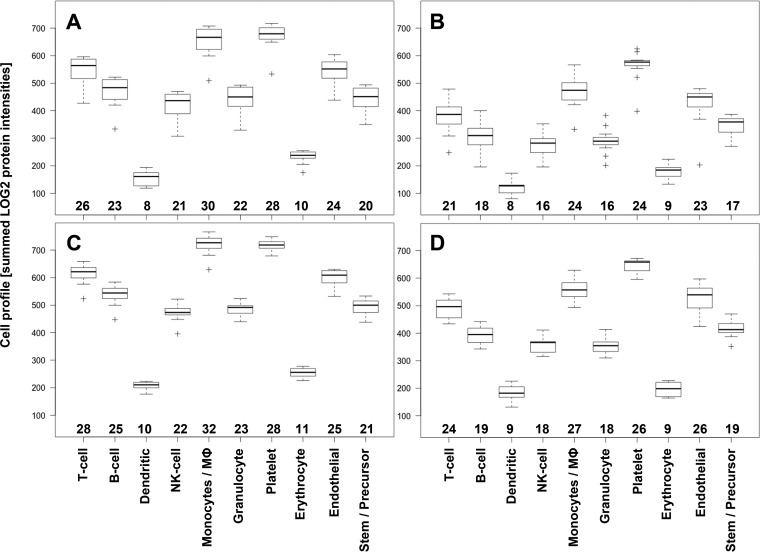
Cells of the vascular system release spherical vesicles, called microparticles, in the size range of 0.1-1 μm induced by a variety of stress factors resulting in variable concentrations between health and disease. Furthermore, microparticles have intercellular communication/signaling properties and interfere with inflammation and coagulation pathways. Today's most used analytical technology for microparticle characterization, flow cytometry, is lacking sensitivity and specificity, which might have led to the publication of contradicting results in the past.We propose the use of nano-liquid chromatography two-stage mass spectrometry as a nonbiased tool for quantitative MP proteome analysis.For this, we developed an improved microparticle isolation protocol and quantified the microparticle protein composition of twelve healthy volunteers with a label-free, data-dependent and independent proteomics approach on a quadrupole orbitrap instrument.Using aliquots of 250 μl platelet-free plasma from one individual donor, we achieved excellent reproducibility with an interassay coefficient of variation of 2.7 ± 1.7% (mean ± 1 standard deviation) on individual peptide intensities across 27 acquisitions performed over a period of 3.5 months. We show that the microparticle proteome between twelve healthy volunteers were remarkably similar, and that it is clearly distinguishable from whole cell and platelet lysates. We propose the use of the proteome profile shown in this work as a quality criterion for microparticle purity in proteomics studies. Furthermore, one freeze thaw cycle damaged the microparticle integrity, articulated by a loss of cytoplasm proteins, encompassing a specific set of proteins involved in regulating dynamic structures of the cytoskeleton, and thrombin activation leading to MP clotting. On the other hand, plasma membrane protein composition was unaffected. Finally, we show that multiplexed data-independent acquisition can be used for relative quantification of target proteins using Skyline software. Mass spectrometry data are available via ProteomeXchange (identifier PXD003935) and panoramaweb.org (https://panoramaweb.org/labkey/N1OHMk.url).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
Disclosures: None.
Figures
References
-
- van der Pol E., Böing A. N., Gool E. L., and Nieuwland R. (2016) Recent developments in the nomenclature, presence, isolation, detection and clinical impact of extracellular vesicles. J. Thromb. Haemost 14, 48–56 - PubMed
-
- Boulanger C. M., Amabile N., and Tedgui A. (2006) Circulating microparticles: a potential prognostic marker for atherosclerotic vascular disease. Hypertension 48, 180–186 - PubMed
-
- Meziani F., Tesse A., and Andriantsitohaina R. (2008) Microparticles are vectors of paradoxical information in vascular cells including the endothelium: role in health and diseases. Pharmacol. Rep. 60, 75–84 - PubMed
-
- Chironi G. N., Boulanger C. M., Simon A., George F. D., Freyssinet J. M., and Tegui A. (2009) Endothelial microperticles in diseases. Cell Tissue Res. 335, 143–151 - PubMed
-
- Little K. M., Smalley M., Harthun N. L., and Ley K. (2010) The plasma microvesicle proteome. Semin. Thromb. Hemost 36, 845–856 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
